The general workflow for single-cell microbial metabolic analysis with genetic identification by OPTIR-FISH is summarized in Figure 2. The representative results demonstrating the single-cell metabolic imaging capability of OPTIR are shown in Figure 3. This example used E. coli cells incubated with 12C- or 13C-glucose for 24 h. The incorporation of 13C into proteins has been shown to cause a significant red shift of protein amides I and II (Figure 3A)17,18. Therefore, these two key wavenumbers representing the 12C-protein (1656 cm-1, normal amide I band) and 13C-protein (1612 cm-1, shifted amide I band) were selected, and OPTIR images at these two wavenumbers were acquired. The corresponding bright-field images are also shown to demonstrate the morphology of the cells (Figure 3B,E). Rod-shaped bacteria can be clearly resolved at the single cell level in both bright field and OPTIR images, confirming the high spatial resolution of the OPTIR technique (Figure 3B–G). For cells incubated with 12C-glucose (Figure 3B–D), a higher intensity at 1656 cm-1 was observed, while a higher 1612 cm-1 intensity was observed for cells incubated with 13C-glucose (Figure 3E–G). This peak shift indicates the incorporation of heavier carbon isotopes into protein mass17. The evaluation was extended to assess OPTIR-FISH's capability in discriminating bacterial taxa and their metabolic activities in multi-species samples. We artificially mixed two bacterial species that are prevalent in the human gut microbiome: E. coli and B. thetaiotaomicron (B. theta). E. coli cells incubated with 13C-glucose were hybridized with a Gam42a-Cy5 probe, targeting the 23S rRNA of Gammaproteobacteria10. B. theta cells incubated with 12C-glucose were hybridized with the Bac303-Cy3 probe, targeting the 16S rRNA of most Bacteroidaceae species11. It is very hard to differentiate the two species based on the bright-field images since they are both rod-shaped (Figure 4A). Thus, multi-color fluorescence imaging of hybridized probes was essential to assign a taxonomic identity to each analyzed bacterial cell (Figure 4B). Upon subtracting OPTIR images acquired at 1612 cm-1 from that of 1656 cm-1, we noticed that a segment of cells in this bi-species sample displayed positive subtraction values (Figure 4C), indicating the incorporation of 13C isotopes. Based on the culture conditions, we assigned the cells with positive subtraction value to be E. coli, and negative subtraction value to be B. theta. The assignment of bacterial species is confirmed by the fluorescence imaging results: the cells producing positive subtraction values produce Cy5 contrast, which is specific to E. coli, and the cells producing negative subtraction values produce B. theta-specific Cy3 contrasts. Then, the isotopic replacement ratio was calculated, in this case, newly synthesized 13C-protein from 13C-glucose (Figure 4D). As expected, a significant difference in isotopic replacement ratio between the two bacterial species (pairwise t-test, p = 9.74 x 10-33) was observed. This dataset underscores the feasibility of applying the OPTIR-FISH platform to study metabolism in a complex environment where multiple species of samples are present.
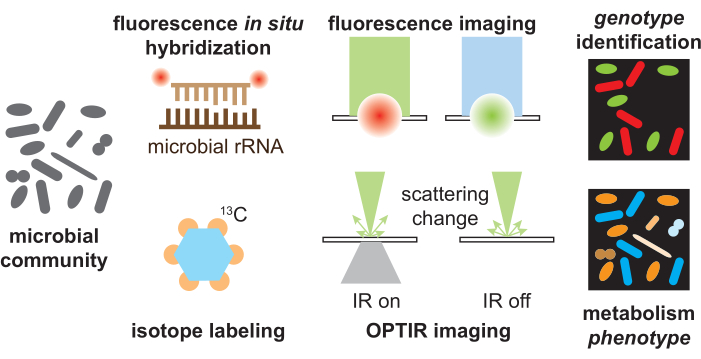
Figure 1: Schematic for the OPTIR-FISH platform for simultaneous species identification and metabolism analysis. Different members of the microbial community could be identified by specific binding of the microbial ribosomal RNA probes through fluorescence in situ hybridization and detected by fluorescence imaging of the attached fluorophores. Metabolic activity could be analyzed by culturing with isotopic substrates and detected by OPTIR imaging. OPTIR imaging detects the scattering change induced by the IR absorption, and OPTIR is inherently compatible with widely used fluorescent tools. Please click here to view a larger version of this figure.
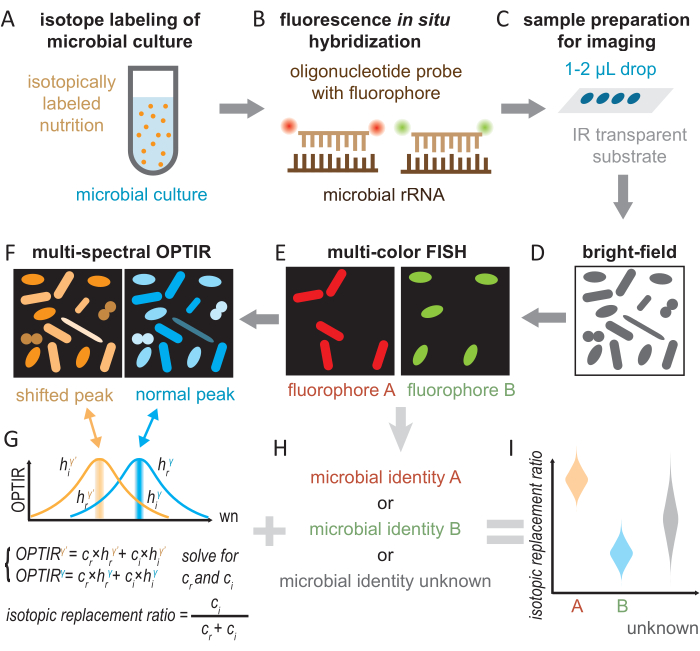
Figure 2: General workflow for single-cell analysis of metabolic activity with genetic identification by OPTIR-FISH. (A) Bacterial cells were cultured with isotopically labeled substrates. (B) Fluorescence in situ hybridization (FISH) was then performed. (C) Sample preparation on IR-transparent slide. (D) Bright-field images identify the regions with optimal cell density. (E,H) Multi-color fluorescence imaging of different FISH fluorophores reveals the genetic identity of individual cells as specified by FISH probes or remains unknown. (F,G) Subsequent multi-spectral OPTIR imaging at the normal and shifted peak reveals the isotopic incorporation into the microbial biomass. Referencing the OPTIR spectra for unlabeled and fully labeled samples (coefficient h), the relative contribution of regular (cr) and isotopic (ci) bio components could be quantified. Protein synthesis from metabolized isotopically-labeled substrates was quantified using the "isotopic replacement ratio". (I) By OPTIR-FISH, a high-throughput analysis of the metabolic activity of microbial species in a complex population can be obtained. Please click here to view a larger version of this figure.
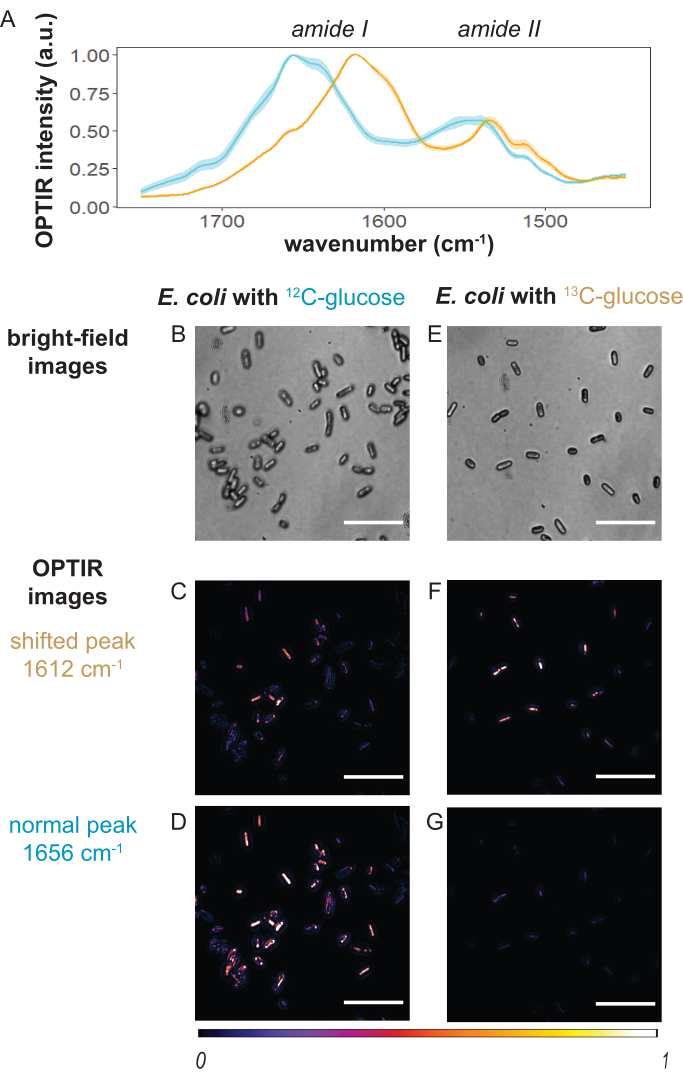
Figure 3: Representative OPTIR spectra and imaging results. (A) OPTIR spectra for E. coli cells cultured with 12C-glucose (cyan) and 13C-glucose (orange) covering the amide I and amide II region. A clear red-shifted amide I peak (1656 cm-1 to 1612 cm-1) is observed for 13C-glucose cultured cells. Representative bright-field and OPTIR images of E. coli cells at normal amide I and shifted amide I band under (B–D) 12C-glucose and (E–G) 13C-glucose culturing conditions. Scale bars: 10 µm. This figure was modified with permission from Bai et al.9. Please click here to view a larger version of this figure.
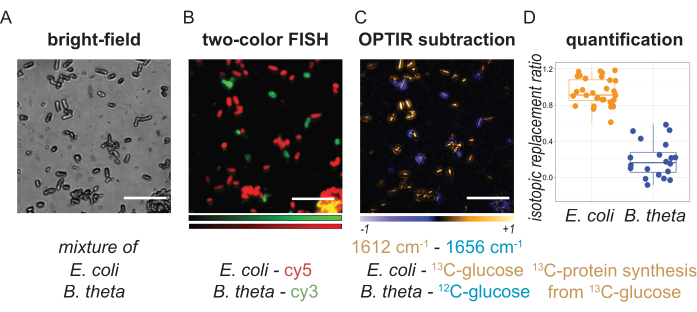
Figure 4: Representative images and quantification results from bacterial mixtures. E. coli cells were incubated with 13C-glucose, followed by hybridization with oligonucleotide probe Gam42a-Cy5. B. theta cells were incubated with 12C-glucose, followed by hybridization with oligonucleotide probe Bac303-Cy3. (A) Bright-field image shows the morphology of the cell mixture. (B) The two-color fluorescence image shows the distribution of E. coli (red) and B. theta (green). (C) OPTIR subtraction result (1612 cm-1-1656 cm-1) shows the protein metabolism of the bacteria cells. (D) Quantification of isotopic replacement ratio representing newly synthesized 13C-protein from 13C-glucose. (Pairwise t-test: p = 9.74 x 10-33). Scale bars: 10 µm. This figure was modified with permission from Bai et al.9. Please click here to view a larger version of this figure.
