Preparation of Single-cell Suspension of Mouse Thymic Epithelial Cells and Staining of Intracellular Molecules for Flow Cytometric Analysis
Summary
We describe protocols for the preparation of single-cell suspension of mouse thymic epithelial cells and the staining of intracellular molecules for flow cytometric analysis.
Abstract
Thymic epithelial cells (TECs) play an essential role in promoting the development and repertoire selection of T cells. Cortical TECs (cTECs) in the thymic cortex induce early T cell development and positive selection of cortical thymocytes. In contrast, medullary TECs (mTECs) in the thymic medulla attract positively selected thymocytes from the cortex and establish self-tolerance in T cells. A variety of molecules, including DLL4 and beta5t expressed in cTECs, as well as Aire and CCL21 expressed in mTECs, contribute to thymus function supporting T cell development and selection. Flow cytometric analysis of functionally relevant molecules in cTECs and mTECs is useful to improve our understanding of the biology of TECs, even though current methods for the preparation of single-cell suspensions of TECs can retrieve only a small fraction of TECs (approximately 1% for cTECs and approximately 10% for mTECs) from young adult mouse thymus. Because many of these functionally relevant molecules in TECs are localized within the cells, we describe our protocols for the preparation of single-cell suspension of mouse TECs and the staining of intracellular molecules for flow cytometric analysis.
Introduction
The thymus is an epithelial mesenchymal organ that promotes the development and repertoire selection of T lymphocytes. In the cortical and medullary microenvironments within the thymus, thymic epithelial cells (TECs) play a crucial role in directing T cell development and selection. It is well accepted that cortical TECs (cTECs) in the thymic cortex primarily contribute to the induction of early T cell development and positive selection of newly generated T-cell antigen-receptor (TCR)-expressing thymocytes. In contrast, medullary TECs (mTECs) in the thymic medulla are essential for the attraction of positively selected thymocytes from the cortex and the establishment of central self-tolerance by eliminating self-reactive T cells and by generating regulatory T cells1,2. These functions of TECs in supporting T cell development and selection are mediated through a variety of molecules expressed in cTECs and mTECs.
These include Delta-like ligand 4 (DLL4), important for T-cell-lineage specification of lymphoid progenitors)3,4, beta5t (also known as Psmb11, important for positive selection of CD8 T cells)5,6, and Prss16 (also known as thymus-specific serine protease, or TSSP, important for positive selection of CD4 T cells)7,8 expressed in cTECs, and Autoimmune regulator (Aire, important for promiscuous expression of self-genes)9,10, Fez family zinc finger 2 (Fezf2, important for promiscuous expression of Aire-independent self-genes and the generation of thymic tuft cells)11,12,13,14, and C-C motif chemokine ligand 21 (CCL21, important for medullary migration and accumulation of positively selected thymocytes)14,15,16 expressed in mTECs. Flow cytometry provides a means for quantitatively detecting these functionally significant molecules in TECs, facilitating the assessment of both the development and function of cTECs and mTECs. Since many of the molecules in TECs are primarily expressed within the cells, such as in the cytoplasm (e.g., proteasome component beta5t) and the nucleus (e.g., transcriptional regulator Aire), it is advantageous to quantitatively analyze intracellular molecules in cTECs and mTECs using flow cytometry.
TECs are rare in the thymus. A female mouse at 5 weeks old has approximately 1 × 106 cTECs and approximately 2.5 × 106 mTECs, whereas the thymic cortex contains approximately 3 × 108 CD4+CD8+ thymocytes and the thymic medulla contains approximately 5 × 107 CD4+CD8– and CD4–CD8+ thymocytes17. Therefore, the vast majority (>99%) of cells in the thymus are hematopoietic cells, including thymocytes, rather than TECs, in young adult mice. Moreover, TECs are tightly associated with neighboring cells (e.g., thymocytes) and architectural extracellular components (e.g., collagen) within the thymus, so the liberation of TECs into cell suspension requires protease digestion of the thymus. Conventional methods for enzymatic digestion yield only a small fraction of TECs; approximately 1% (1 × 104) and 10% (2 × 105) for cTECs and mTECs, respectively, from 5-week-old mouse thymus17,18,19. The low efficiency of TEC liberation from the thymus is likely due to the susceptibility of TECs to proteolytic enzymes and mechanical stress and the tight association of TECs with neighboring cells, particularly between cTECs and CD4+CD8+ thymocytes20. In this regard, the results of single-cell analysis of mouse TECs, including flow cytometric analysis and single-cell RNA-sequencing analysis, based on current enzymatic digestion-mediated cell preparation methods are representative of only a small portion of TECs and therefore, are far from being comprehensive. Nevertheless, despite such limitations in the enzymatic retrieval of TECs, flow cytometric analysis of mouse TECs in single-cell suspension is still useful for the quantitative analysis of functionally relevant molecules in cTECs and mTECs. In cell suspension, TECs have been defined as cells positive for epithelial surface molecule EpCAM (CD326) and negative for hematopoietic surface molecule CD45. Within EpCAM+CD45– TECs, cell-surface expression levels of Ly51 molecules (also known as BP-1 and CD249) and Ulex Europaeus agglutinin I (UEA1) binding capacity (primarily binding to glycoproteins and glycolipids containing α-linked fucose) have been used to phenotypically mark cTECs (Ly51highUEA1low) and mTECs (Ly51lowUEA1high). Here we describe our experimental protocols for the conventional preparation of single-cell suspension of mouse TECs and the staining of intracellular molecules for flow cytometric analysis, focusing on the quantitative analysis of intracellular molecules beta5t, CCL21, and Aire in freshly prepared mouse TECs.
Protocol
All mouse experiments were performed under the approval of the Animal Care and Use Committee of the National Cancer Institute (ASP21-431, ASP21-432, and EIB-076-3).
1. Preparation of thymus cell suspension
NOTE: Mouse ontogeny affects the efficiency of enzymatic liberation of TECs from the thymus. Three methods depending on mouse age are described as follows.
- For postnatal mice more than 3 weeks old
- Harvest the thymus (two thymus lobes) from one mouse. Place the thymus in 5 mL of RPMI1640 containing 10 mM HEPES in a 60 mm plastic dish on ice.
- Remove blood, non-thymus tissues, and fat with curved fine-tip tweezers under a dissecting microscope. Cut the thymus into small pieces (approximately 1-2 mm in size) with dissecting scissors.
- Transfer the thymus pieces into a 15 mL conical tube containing 3 mL of 1 mg/mL Collagenase/Dispase, and 1 U/mL DNase solution in RPMI1640 containing 2% fetal bovine serum (FBS). Incubate in a water bath or a heat block at 37 °C for 20 min.
- Mix gently using a 3 mL plastic transfer pipet. Incubate for another 20 min in case the tissue fragments are still visible.
- Mix gently using a 3 mL plastic transfer pipet. Incubate for another 10 min in case the tissue fragments are still visible.
- Mix gently using a 3 mL plastic transfer pipet. Add 5 mL of 5 mM EDTA in 1x HBSS containing 1% FBS.
- Filter the cell suspension through a 60 µm nylon mesh and into a new 50 mL conical tube. Centrifuge the cell suspension at 350 × g at 4 °C for 5 min.
- Resuspend the cell pellet in 10 mL of FACS buffer (1x HBSS containing 1% bovine serum albumin [BSA] and 0.1% NaN3).
- Store the cells on ice and determine cell numbers using a cell counter.
- For neonatal mice between 1 and 2 weeks old
- Perform step 1.1.1.
- Remove blood and non-thymus tissues with dissecting tweezers under a dissecting microscope. Transfer the thymus into a 1.5 mL plastic tube containing 200 µL of RPMI1640 with 10 mM HEPES and cut the thymus into small pieces (approximately 1-2 mm in size) with dissecting scissors.
- Allow the thymus fragments to settle at the bottom of the tube, then gently remove the suspension with a 200 µL micropipette.
- Transfer the thymus pieces into a 5 mL polypropylene round-bottom tube containing 2 mL of 0.5 mg/mL Collagenase/Dispase and 1 U/mL DNase solution in RPMI1640 containing 2% FBS. Incubate in a water bath or a heat block at 37 °C for 10 min.
- Mix gently using a 3 mL plastic transfer pipet and then, with a 1 mL micropipette fitted with a plastic tip for further homogenization.
- Add 2 mL of 5 mM EDTA in 1x HBSS containing 1% FBS. Filter the cell suspension through a 60 µm nylon mesh and into a 50 mL conical tube. Add 16 mL of RPMI1640 containing 10 mM HEPES.
- Centrifuge the cell suspension 350 × g at 4 °C for 5 min. Resuspend the cell pellet in 10 mL of FACS buffer.
- Store cells on ice and determine cell numbers using a cell counter.
- For embryonic mice and neonatal mice up to 1 week old
- Harvest the thymus (two thymus lobes) from one mouse. Place the thymus in a 60 mm plastic dish and remove blood, non-thymus tissues, and fat with curved fine-tip tweezers under a dissecting microscope.
- Place the thymus in 0.2 mL of RPMI1640 containing 10 mM HEPES in a 1.5 mL microcentrifuge tube. Discard the liquid with a 200 µL micropipette.
- Add 0.5 mL of 0.5 mg/mL Collagenase/Dispase and 1 U/mL DNase solution in RPMI1640 containing 2% FBS. Incubate in a water bath or a heat block at 37 °C for 10 min. Mix gently using a 1 mL micropipette with a plastic tip.
- Add 0.5 mL of 5 mM EDTA in 1x HBSS containing 1% FBS. Filter the cell suspension through a 60 µm nylon mesh and into a new 15 mL conical tube. Add 9 mL of RPMI1640 containing 10 mM HEPES.
- Centrifuge the cell suspension at 350 × g at 4 °C for 5 min. Resuspend the cell pellet in 1-3 mL of FACS buffer.
- Store cells on ice and determine cell numbers using a cell counter.
- Alternative method: Liberase digestion
- Perform steps 1.1.1 and 1.1.2.
- Allow the thymus fragments to settle at the bottom of the tube; then gently remove the suspension with a 200 µL micropipette. Transfer the thymus pieces into a 5 mL tube containing 1 mL of 0.5 U/mL Liberase and 1 U/mL DNase in RPMI1640 containing 10 mM HEPES. Incubate in a water bath or a heat block at 37 °C for 15 min.
- Mix gently using a 1 mL micropipette with a plastic tip. Incubate for another 15 min in case the tissue fragments are still visible.
- Mix gently using a 1 mL micropipette with a plastic tip. Add 1 mL of 5 mM EDTA in 1x HBSS containing 1% FBS.
- Filter the cell suspension through a 60 µm nylon mesh and into a new 50 mL conical tube. Add 18 mL of RPMI1640 containing 10 mM HEPES.
- Centrifuge the cell suspension at 350 × g, 4 °C for 5 min. Resuspend the cells in 10 mL of FACS buffer (1x HBSS containing 1% BSA and 0.1% NaN3).
- Store cells on ice and determine cell numbers using a cell counter.
2. Antibody staining of thymus cells
- Place 16 × 106 cells in a 5 mL polystyrene round-bottom tube. Centrifuge the cells at 350 × g at 4 °C for 5 min. Discard the supernatant.
- Add 10 µL of working concentration of anti-Fc receptor monoclonal antibody (mAb, clone 2.4G2) in FACS buffer to block nonspecific binding of staining IgG to cell-surface Fc receptor. Incubate at 4 °C for 5 min.
- Add 40 µL each of working concentrations of anti-CD45 mAb (hematopoietic cell-specific; e.g., BV421-labeled), anti-EpCAM mAb (epithelial cell-specific; e.g., PE-Cy7-labeled), anti-Ly51 mAb (cTEC-detecting; e.g., AF647-labeled), and Ulex europaeus agglutinin I (UEA-1) (mTEC-detecting; e.g., AF594-labeled) in FACS buffer. In parallel, prepare tubes for single-color stained cells and unstained cells for setting up a flow cytometer.
- Incubate at 4 °C for 45 min. Mix well every 10 min. Add 2 mL of FACS buffer.
- Centrifuge cells 350 × g at 4 °C for 5 min. Discard the supernatant.
- Add 1 µL of Ghost Dye Violet 510 (to distinguish viable cells from dead cells in flow cytometric analysis of fixed and permeabilized cells). Use undiluted Ghost Dye reagent; mix well. Incubate at 4 °C for 30 min; mix well every 10 min.
- Add 2 mL of FACS buffer. Centrifuge cells 350 × g at 4 °C for 5 min. Discard the supernatant.
- Fix the cells by adding 1 mL of 2% paraformaldehyde (PFA) in PBS and incubating at room temperature for 10 min.
- Add 3.5 mL of PBS to the tube and centrifuge cells 700 × g at 4 °C for 8 min. Discard the supernatant.
- Permeabilize the cells by adding 1 mL of Intracellular Fixation/Permeabilization Buffer and incubating at 4 °C for 20 min.
- Centrifuge the cells 700 × g at 4 °C for 8 min. Discard the supernatant.
- Add 1 mL of 1x Permeabilization Buffer. Centrifuge the cells at 700 × g at 4 °C for 8 min. Discard the supernatant.
- Add 100 µL each of working concentrations of anti-beta5t antibody or anti-CCL21 antibody, or 40 µL of working concentrations of anti-Aire antibody (e.g., AF488-labeled), in 1x permeabilization buffer, mix well, and incubate for 60 min at room temperature.
- Add 1 mL of 1x Permeabilization Buffer. Centrifuge the cells at 700 × g at 4 °C for 8 min. Discard the supernatant.
- Repeat step 2.14.
- Add 100 µL of working concentrations of anti-rabbit IgG (e.g., AF555-labeled) and incubate for 30 min at room temperature.
- Add 1 mL of 1x Permeabilization Buffer. Centrifuge the cells at 700 × g at 4 °C for 8 min. Discard the supernatant.
- Resuspend the cells in 200 µL of FACS Buffer and mix well.
- Store the cells in the dark at 4 °C until flow cytometric analysis.
- Filter cells through 60 µm nylon mesh prior to flow cytometric analysis.
Representative Results
The analysis requires a flow cytometer that allows the detection of at least six fluorescence channels (for CD45, EpCAM, Ly51, UEA1, Ghost Dye, and beta5t, CCL21, and/or Aire) as well as forward and side scatter parameters. Representative flow cytometric profiles of Collagenase/Dispase-digested thymus cells from 0-day-old mice and 2-week-old mice are shown in Figure 1 and Figure 2, respectively. In both figures, the dot plot in Panel A shows forward scatter (FSC) intensity and Ghost Dye Violet 510 fluorescence intensity of total particulate signals. More than 90% of particulate signals represent viable cells, as judged by negative staining by the Ghost Dye reagent and by forward scatter reflecting cell size. Panel B shows cell surface fluorescence signals of viable thymus cells for CD45, a molecule expressed by hematopoietic cells, and EpCAM, a molecule expressed by epithelial cells. A small but distinctive cell population is CD45– EpCAM+, therefore representing TECs.
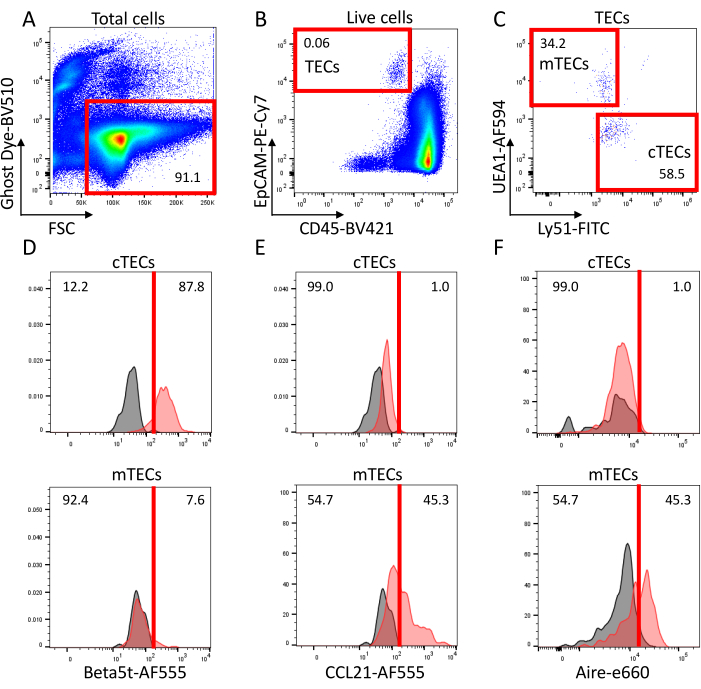
Figure 1: Representative flow cytometric profiles of Collagenase/Dispase-digested thymus cells from 0-day-old wild-type C57BL/6-background mice. (A) Forward scatter intensity and Ghost Dye Violet 510 fluorescence intensity of total particulate signals. Red box indicates the signals of viable cells. (B) Fluorescence signals of viable cells (red box in panel A) for CD45 and EpCAM. Red box indicates CD45– EpCAM+ TECs. (C) Ly51 and UEA1 cell-surface staining signals in TECs (red box in panel B) reveal two major populations: Ly51high UEA1low cells enriched with cTECs and Ly51low UEA1high cells enriched with mTECs. (D-F) Detection of (D) beta5t, (E) CCL21, and (F) Aire in cTECs (top panels) and mTECs (bottom panels) as defined in panel C. Gray histograms show control profiles stained with (D,E) secondary antibody only and (F) stained using cells from Aire-deficient mice. Abbreviations: FSC = forward scatter; TECs = Thymic epithelial cells; cTECs = cortical TECs; mTECs = medullary TECs. Please click here to view a larger version of this figure.
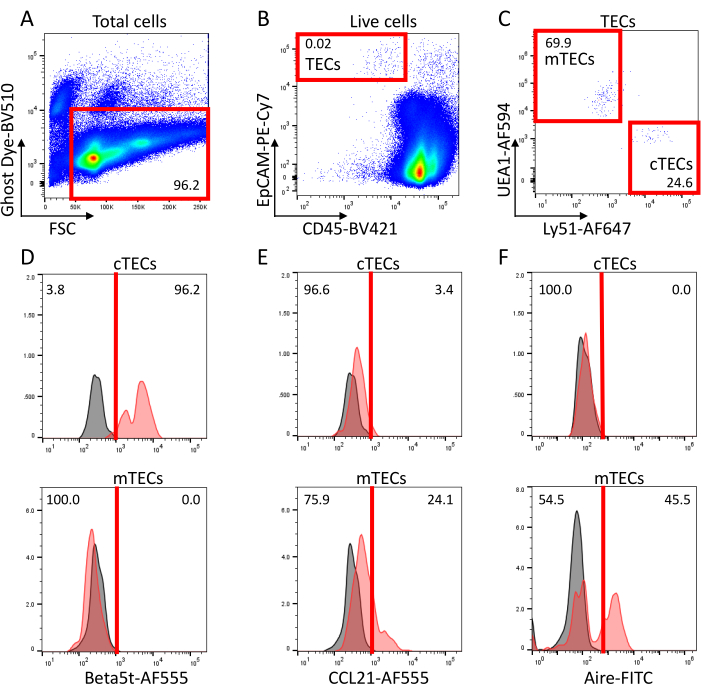
Figure 2: Representative flow cytometric profiles of Collagenase/Dispase-digested thymus cells from 2-week-old wild-type C57BL/6-background mice. (A) Forward scatter intensity and Ghost Dye Violet 510 fluorescence intensity of total particulate signals. Red box indicates the signals of viable cells. (B) Fluorescence signals of viable cells (red box in panel A) for CD45 and EpCAM. Red box indicates CD45– EpCAM+ TECs. (C) Ly51 and UEA1 cell-surface staining signals in TECs (red box in panel B) reveal two major populations: Ly51high UEA1low cells enriched with cTECs and Ly51low UEA1high cells enriched with mTECs. (D-F) Detection of (D) beta5t, (E) CCL21, and (F) Aire in cTECs (top panels) and mTECs (bottom panels) as defined in panel C. Gray histograms show control profiles stained with (D,E) secondary antibody only and stained with (F) FITC-labeled isotype-matched IgG. Abbreviations: FSC = forward scatter; TECs = Thymic epithelial cells; cTECs = cortical TECs; mTECs = medullary TECs; FITC = fluorescein isothiocyanate. Please click here to view a larger version of this figure.
Ly51 and UEA1 cell-surface staining signals as shown in Panel C reveal two major populations in TECs; a population relatively high in Ly51 and low in UEA1 enriched with cTECs, and another relatively low in Ly51 and high in UEA1 enriched with mTECs21,22,23,24,25. During embryonic and neonatal stages, including 0-day-old, the frequency of cTECs is larger than that of mTECs. In postnatal stages, the frequency of mTECs dominates mainly because the efficiency of cTEC liberation from the thymus becomes extremely low (approximately 1% of cTECs and 10% of mTECs can be retrieved into a single cell suspension)17.
Fixation and permeabilization of cells allow the detection of intracellular molecules within TECs. Panel D shows beta5t expression detected in the majority of cTECs but not in mTECs; panel E shows the detection of CCL21 in a fraction of mTECs but not in cTECs, and panel F shows Aire expressed in a subpopulation of mTECs.
Additional flow cytometric profiles of intracellular molecules in mouse TECs may be found in published articles; for beta5t6,20, CCL2114,16,26, and Aire27,28, as well as for additional intracellular molecules, including keratin22,29 and Fezf211,30.
Discussion
Collagenase, with or without Dispase, is widely used to digest mouse thymus to prepare TECs4,5,9,18,21. Liberase, which contains a blend of proteases including collagenase, is also used to digest the thymus to prepare TECs17,22. The use of Collagenase and Liberase gives essentially equivalent yields and quality of cTECs and mTECs from mouse thymus, although minor differences may appear depending on the product batch of the enzymes. Trypsin is also used for efficient thymus digestion21,23, although it has been shown that many cell-surface molecules, including EpCAM and CD80 expressed by TECs, are susceptible to trypsin digestion21. For enzymatic digestion of the thymus, DNase is often added to reduce viscosity owing to DNA release from damaged cells17,21.
Enzymatic digestion of the thymus is essential for the liberation of TECs from the thymus. The thymus should be sufficiently digested until tissue fragments become invisible and the cell suspension appears homogeneous. However, digestion should not be left too long because excessive enzymatic treatment damages cells and diminishes cell viability. In every experiment, it is important to pay careful attention to the incubation period and the pipetting strength during the thymus digestion. It is also important to be aware that current enzymatic digestion technology allows the liberation of only a small fraction of TECs, as detailed in the Introduction and as reported previously17,18,19.
Because TECs are rare in total thymus cell suspension, flow cytometric analysis of TECs using total thymus cell suspension requires the staining of many cells, typically 8 × 106 to 16 × 106 cells per measurement.
TECs derived from enzyme-digested thymus cells may be enriched using anti-CD45 magnetic beads22 and/or Percoll density gradient centrifugation31. These enrichment procedures are timesaving especially when TECs are subsequently purified over a flow cytometric cell sorter. It should also be noted that the anti-CD45 enrichment would remove a subpopulation of TECs that are tightly associated with CD45-expressing thymocytes20 and the Percoll centrifugation might remove an unexpectedly dense subpopulation of TECs.
Regarding the fixation of TECs, it is known that PFA treatment shrinks cells and reduces the forward scatter intensity in flow cytometric analysis. A gentle pipetting of cells immediately after the addition of 2% PFA and the subsequent addition of PBS may help prevent excessive cell shrinkage.
For flow cytometric analysis, given the infrequent occurrence of TECs within the thymus, their delineation necessitates a distinct approach compared to thymocytes or splenocytes. Initial classification is grounded in the utilization of CD45, a hematopoietic pan-marker ubiquitous on thymocytes and other hematopoietic cells, including dendritic cells and macrophages. Conversely, TECs, being non-hematopoietic in origin, lack CD45 expression. Instead, TECs express EpCAM on their surface, hence, allowing a straightforward gating strategy of CD45-negative and EpCAM-positive to discern TECs. In our flow cytometric analysis, we commence by defining this population and subsequently ensure appropriate scaling of forward and side scatter parameters, given the substantial size discrepancy between TECs and thymocytes, and given the large size and high autofluorescence of TECs.
Subsequently, attention is directed towards the evaluation of cTEC- and mTEC-markers within the CD45-negative EPCAM-positive population. Owing to the diminutive size of this population, this step becomes imperative for compensating cTECs and mTECs. Examination of an all-stained panel (e.g., cTEC-marker Ly51 and mTEC-marker UEA-1 along with TEC-defining CD45 and EpCAM) further ensures the adequacy of scatter scaling. During flow cytometric setup, our primary objective revolves around discerning positive and negative populations for each antibody.
In the co-staining of Aire, beta5t, and CCL21, distinct populations within TEC subsets are identified. Given Aire's selective expression by mTECs, we focus on the four-color stain of CD45-negative/EPCAM-positive/UEA-1-positive/Ly51-negative cells. Here, UEA-1-positive cells are utilized to establish the negative control for Aire, facilitating clear identification of Aire-positive cells. A similar approach is adopted for CCL21, which is expressed within mTECs. Conversely, for beta5t, expressed exclusively by cTECs, a comparable gating strategy is applied, with emphasis on Ly51-positive cells within the four-color stain, to ascertain background levels for beta5t. Subsequent analysis of samples stained with anti-beta5t facilitates the identification of a positive population.
Anti-beta5t antibody and anti-CCL21 antibody are rabbit antibodies, so these antibodies should not be mixed without prior fluorescence labeling. Working concentrations of antibodies should be predetermined by performing flow cytometric analysis of control cells (e.g., TECs from wild-type and knockout mice) stained with serial dilutions of antibodies.
In flow cytometric analysis, cTECs and mTECs are frequently defined by the cell-surface expression of Ly51 or CD205 for cTECs and of UEA1 and CD80 for mTECs. However, it should be kept in mind that cTECs and mTECs must be eventually defined by their microanatomical localization in the thymus. In the embryonic thymus, emerging evidence has suggested that the expression of Ly51 and CD205 is not limited to cTECs but extends to other populations of TECs, including cTEC/mTEC-bipotent progenitors and immature mTECs32,33. Additionally, Ly51 expression in cTECs remains low during embryogenesis and perinatal period in comparison with postnatal period33, as also shown in Figure 1 and Figure 2.
In conclusion, flow cytometric analysis of enzymatically liberated single-cell suspension of TECs offers a powerful technique for the quantitative analysis of TECs and their subpopulations at single-cell resolution, despite the limited efficiency of the liberation of TECs. In particular, flow cytometric analysis of intracellular molecules in single-cell TEC suspension is useful for studying the expression of a variety of molecules important for the development and function of TECs and their subpopulations.
Declarações
The authors have nothing to disclose.
Acknowledgements
We thank Dr. Izumi Ohigashi for reading the manuscript. This work was supported by the Intramural Research Program ZIA BC 011806 of the National Institutes of Health, the National Cancer Institute, and the Center for Cancer Research.
Materials
| Collagenase/Dispase | Roche | 11097113001 | |
| Liberase TM | Roche | 5401127001 | |
| Foxp3 / Transcription Factor Staining Buffer Set | eBioscience | 00-5523-00 | |
| Ghost Dye Violet 510 | Tonbo | 13-0870-T500 | |
| BV421 anti-mouse CD45 monoclonal antibody, Clone 30-F11 | BD Horizon | 563890 | |
| Alexa Fluor 647 anti-mouse Ly51 monoclonal antibody, Clone 6C3 | BioLegend | 108312 | |
| PE/Cy7 anti-mouse EpCAM/CD326 monoclonal antibody, Clone G8.8 | BioLegend | 118216 | |
| DyLight 594 Ulex Europaeus Agglutinin I (UEA1) | Vector Laboratories | DL-1067-1 | |
| Alexa Fluor 488 anti-mouse Aire monoclonal antibody, Clone 5H12 | eBioscience | 53-5934-82 | |
| Anti-mouse Psmb11/beta5t rabbit monoclonal antibody, Clone CPTC-PSMB11(mouse)-1 | NIH NCI | CPTC-PSMB11(mouse)-1 | https://proteomics.cancer.gov/antibody-portal |
| Anti-mouse CCL21/exodus 2 rabbit polyclonal antibody | Bio-Rad | AAM27 | |
| Alexa Fluor 555 anti-rabbit IgG heavy chain goat recombinant antibody | Invitrogen | A27039 | |
| Anti-Mouse CD32/CD16 – Purified (Fc Block Antibody) | Leinco Technologies | C247 | |
| Nylon Mesh 60 Micron | Component Supply | W31436 | |
| Equipment | |||
| Thermomixer | Eppendorf | Heat block | |
| Cellometer Auto 2000 Cell Viability counter | Nexcelom | Viable cell counting | |
| LSRFortessa Cell Analyzer | BD Biosciences | Flow cytometer | Five lasers (355, 408, 488, 595, and 637 nm) |
| FACSDiva software version 9.0.1 | BD Biosciences | Flow cytometric analysis | |
| FlowJo 10.9.0 | BD Biosciences | Flow cytometric data analysis | |
| Antibody | Dilution rate (Working concentration) | Dilution Buffer | |
| Anti-Mouse CD32/CD16 – Purified (Fc Block Antibody) | 1:40 | FACS Buffer | |
| BV421 anti-mouse CD45 monoclonal antibody | 1:25 | FACS Buffer | |
| PE/Cy7 anti-mouse EpCAM/CD326 monoclonal antibody | 1:100 | FACS Buffer | |
| Alexa Fluor 647 anti-mouse Ly51 monoclonal antibody | 1:100 | FACS Buffer | |
| DyLight 594 Ulex Europaeus Agglutinin I (UEA1) | 1:400 | FACS Buffer | |
| Anti-mouse CCL21/exodus 2 rabbit polyclonal antibody | 1:200 | 1x Permeabilization Buffer | |
| Anti-mouse Psmb11/beta5t rabbit monoclonal antibody, Clone CPTC-PSMB11(mouse)-1 | 1:1000 | 1x Permeabilization Buffer | |
| Alexa Fluor 555 anti-rabbit IgG heavy chain goat recombinant antibody | 1:400 | 1x Permeabilization Buffer | |
| Buffer | Purpose | ||
| RPMI1640 containing 10 mM HEPES | For cell preparation | ||
| RPMI1640 containing 2% FBS | For cell preparation | ||
| FACS buffer (1x HBSS containing 1% BSA and 0.1% sodium azide) | For cell preparation and antibody staining | ||
| 1 mg/ml Collagenase/Dispase and 1U/ml DNase solution in RPMI1640 containing 2% FBS | For enzyme digestion | ||
| 2% PFA in PBS | For cell fixation | ||
| Fixation/Permeabilization Buffer (1 part of Fixation/Permeabilization Concentrate with 3 parts of Fixation/Permeabilization Diluent) | For cell fixation | ||
| 1x Permeabilization Buffer (1 part 10X concentrate with 9 parts distilled water) | For intercellular antibody staining |
Referências
- Klein, L., Kyewski, B., Allen, P. M., Hogquist, K. A. Positive and negative selection of the T cell repertoire: what thymocytes see (and don’t see). Nat Rev Immunol. 14 (6), 377-391 (2014).
- Takahama, Y., Ohigashi, I., Baik, S., Anderson, G. Generation of diversity in thymic epithelial cells. Nat Rev Immunol. 17 (5), 295-305 (2017).
- Hozumi, K., et al. Delta-like 4 is indispensable in thymic environment specific for T cell development. J Exp Med. 205 (11), 2507-2513 (2008).
- Koch, U., et al. Delta-like 4 is the essential, nonredundant ligand for Notch1 during thymic T cell lineage commitment. J Exp Med. 205 (11), 2515-2523 (2008).
- Murata, S., et al. Regulation of CD8+ T cell development by thymus-specific proteasomes. Science. 316 (5829), 1349-1353 (2007).
- Ripen, A. M., Nitta, T., Murata, S., Tanaka, K., Takahama, Y. Ontogeny of thymic cortical epithelial cells expressing the thymoproteasome subunit β5t. Eur J Immunol. 41 (5), 1278-1287 (2011).
- Gommeaux, J., et al. Thymus-specific serine protease regulates positive selection of a subset of CD4+ thymocytes. Eur J Immunol. 39 (4), 956-964 (2009).
- Viret, C., et al. Thymus-specific serine protease contributes to the diversification of the functional endogenous CD4 T cell receptor repertoire. J Exp Med. 208 (1), 3-11 (2011).
- Anderson, M. S., et al. Projection of an immunological self shadow within the thymus by the aire protein. Science. 298 (5597), 1395-1401 (2002).
- Mathis, D., Benoist, C. Aire. Annu Rev Immunol. 27, 287-312 (2009).
- Takaba, H., et al. Fezf2 orchestrates a thymic program of self-antigen expression for immune tolerance. Cell. 163 (4), 975-987 (2015).
- Takaba, H., Takayanagi, H. The mechanisms of T cell selection in the thymus. Trends Immunol. 38 (11), 805-816 (2017).
- Lammers, S., et al. Ehf and Fezf2 regulate late medullary thymic epithelial cell and thymic tuft cell development. Front Immunol. 14, 1277365 (2024).
- Ushio, A., et al. Functionally diverse thymic medullary epithelial cells interplay to direct central tolerance. Cell Rep. 43 (4), 114072 (2024).
- Kozai, M., et al. Essential role of CCL21 in establishment of central self-tolerance in T cells. J Exp Med. 214 (7), 1925-1935 (2017).
- Lkhagvasuren, E., Sakata, M., Ohigashi, I., Takahama, Y. Lymphotoxin β receptor regulates the development of CCL21-expressing subset of postnatal medullary thymic epithelial cells. J Immunol. 190 (10), 5110-5117 (2013).
- Sakata, M., Ohigashi, I., Takahama, Y. Cellularity of thymic epithelial cells in the postnatal mouse. J Immunol. 200 (4), 1382-1388 (2018).
- Hirakawa, M., et al. Fundamental parameters of the developing thymic epithelium in the mouse. Sci Rep. 8 (1), 11095 (2018).
- Venables, T., Griffith, A. V., DeAraujo, A., Petrie, H. T. Dynamic changes in epithelial cell morphology control thymic organ size during atrophy and regeneration. Nat Commun. 10 (1), 4402 (2019).
- Nakagawa, Y., et al. Thymic nurse cells provide microenvironment for secondary T cell receptor α rearrangement in cortical thymocytes. Proc Natl Acad Sci USA. 109 (50), 20572-20577 (2012).
- Gray, D. H., Chidgey, A. P., Boyd, R. L. Analysis of thymic stromal cell populations using flow cytometry. J Immunol Methods. 260 (1-2), 15-28 (2002).
- Seach, N., Wong, K., Hammett, M., Boyd, R. L., Chidgey, A. P. Purified enzymes improve isolation and characterization of the adult thymic epithelium. J Immunol Methods. 385 (1-2), 23-34 (2012).
- Jenkinson, E. J., Anderson, G., Owen, J. J. Studies on T cell maturation on defined thymic stromal cell populations in vitro. J Exp Med. 176 (3), 845-853 (1992).
- Adkins, B., Tidmarsh, G. F., Weissman, I. L. Normal thymic cortical epithelial cells developmentally regulate the expression of a B-lineage transformation-associated antigen. Immunogenetics. 27 (3), 180-186 (1988).
- Farr, A. G., Anderson, S. K. Epithelial heterogeneity in the murine thymus: fucose-specific lectins bind medullary epithelial cells. J Immunol. 134 (5), 2971-2977 (1985).
- Wells, K. L., et al. Combined transient ablation and single-cell RNA-sequencing reveals the development of medullary thymic epithelial cells. eLife. 9, e60188 (2020).
- Hubert, F. X., et al. A specific anti-Aire antibody reveals aire expression is restricted to medullary thymic epithelial cells and not expressed in periphery. J Immunol. 180 (6), 3824-3832 (2008).
- Nishikawa, Y., et al. Temporal lineage tracing of Aire-expressing cells reveals a requirement for Aire in their maturation program. J Immunol. 192 (6), 2585-2592 (2014).
- Jenkinson, W. E., Rossi, S. W., Parnell, S. M., Jenkinson, E. J., Anderson, G. PDGFR alpha-expressing mesenchyme regulates thymus growth and the availability of intrathymic niches. Blood. 109 (3), 954-960 (2007).
- Cosway, E. J., et al. Redefining thymus medulla specialization for central tolerance. J Exp Med. 214 (11), 3183-3195 (2017).
- Farr, A. G., Eisenhardt, D. J., Anderson, S. K. Isolation of murine thymic epithelium and an improved method for its propagation in vitro. Anat Rec. 216 (1), 85-94 (1986).
- Baik, S., Jenkinson, E. J., Lane, P. J., Anderson, G., Jenkinson, W. E. Generation of both cortical and Aire+ medullary thymic epithelial compartments from CD205+ progenitors. Eur J Immunol. 43 (3), 589-594 (2013).
- Ohigashi, I., et al. Developmental conversion of thymocyte-attracting cells into self-antigen-displaying cells in embryonic thymus medulla epithelium. eLife. 12, RP92552 (2024).

.