Supplementation of the C. elegans diet is limited by the ability of the bacterial food source to uptake and incorporate fatty acid into the bacterial membrane. To determine the ability of E. coli OP50 to assimilate various fatty acids into its membranes, OP50 was plated onto media with no supplement, 0.1 mM and 0.3 mM concentrations of stearic acid (18:0), sodium oleate (18:1n-9), and sodium DGLA (20:3n-6). Plates were dried at room temperature for 2 days in the dark, and incubated at 20 °C for 3 days. Bacterial lawns were collected by gently scraping the lawn into water with a flame-sterilized spatula. Bacteria were pelleted by centrifugation, and treated with 2.5% H2SO4 in methanol to produce fatty acid methyl esters, which were analyzed by GC/MS following the methods listed in the Procedure, step 3. The results demonstrate that unsaturated fatty acids (oleate and DGLA) incorporate into OP50 in higher amounts than the saturated fatty acid stearic acid (Figure 1A).
Additionally, L1 stage N2 larvae were grown on the same batch of supplemented plates and harvested after three days growth at 20 °C. Worms were washed off of the plates and fatty acids in total worm preps were analyzed by GC/MS. The change in supplemented fatty acids is graphed in Figure 1B. These studies demonstrate that supplementation of saturated fatty acids does not change the relative amount of saturated fatty acids in worm tissues, while supplementation of unsaturated fatty acids increased the relative amounts of unsaturated fatty acids in C. elegans lipids. Taken together, the data shown in Figure 1A and Figure 1B demonstrate that the relative accumulation of supplemented fatty acids in C. elegans correlates directly with the relative accumulation of fatty acids in the dietary E. coli.
We have previously shown that dietary DGLA causes sterility in C. elegans10. Figure 2 illustrates the dose response of DGLA induction of sterility in C. elegans. The concentration of DGLA in worm lipids in which 50% of the population will be sterile is approximately 12%. Interestingly, the response to DGLA can be altered by genetic mutations in C. elegans. A recent finding is that the insulin growth factor-dependent stress pathways can suppress the DGLA-induced germ cell destruction8. Supplementing the diet of worms containing deleterious mutations in either the daf-2 insulin/IGF receptor, daf-2(e1370), or the daf-16/FOXO transcription factor, daf-16(mu86), illustrates the usefulness of this method to unravel genetic pathways that influence the physiological effects of dietary fats. Synchronized L1 larvae were pipetted onto DGLA supplemented media. After 3-4 days of growth, worms were scored for sterility, as determined by the absence of eggs in the uterus of adult worms. DGLA supplemented daf-2(e1370)mutants were fertile, with little to no induced germ cell loss compared to wild type (N2) worms at both 0.15 mM and 0.3 mM supplementations (Figure 3).In contrast, DGLA supplemented worms with inactive FOXO (daf-16(mu86)) displayed a higher percentage of sterile worms compared to wild type when fed on plates containing 0.15 mM DGLA (Figure 3).
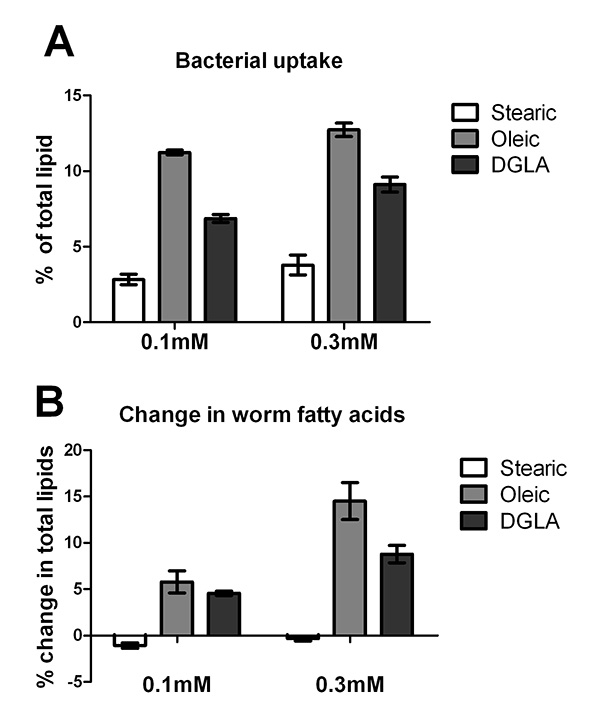
Figure 1. Uptake and incorporation of supplemented fatty acids by E. coli OP50 and C. elegans. A. E. coli OP50 was grown on plates containing 0.1 mM or 0.3 mM stearic acid, sodium oleate, or sodium DGLA as well as un-supplemented plates. After five days of growth on plates at 20 °C, E. coli were harvested and fatty acid methyl esters were generated for analysis by GC/MS. Because OP50 does not produce oleic acid or DGLA, and produces only trace amounts of stearic acid, the percentage of each supplemented fatty acid in the E. coli lipids reveals the ability of OP50 to incorporate the supplemented fatty acid. Error bars are SD. B. Change in C. elegans fatty acids in young adults grown for three days, starting at L1 stage, on E. coli plates containing 0.1 mM or 0.3 mM stearic acid, sodium oleate, or DGLA. The values for change in stearic acid and DGLA were obtained by subtracting the relative amount of 18:0 or 20:3 in worms grown on supplemented plates from those of worms grown on unsupplmented plates. To monitor uptake of oleic acid, the sum of oleic acid plus downstream C20 PUFAs (20:3, 20:4n-6, 20:4n-3, and 20:5) were calculated in supplemented and unsupplemented plates, because incorporated oleic acid is further desaturated and elongated. Error bars are SD. Click here to view larger image.
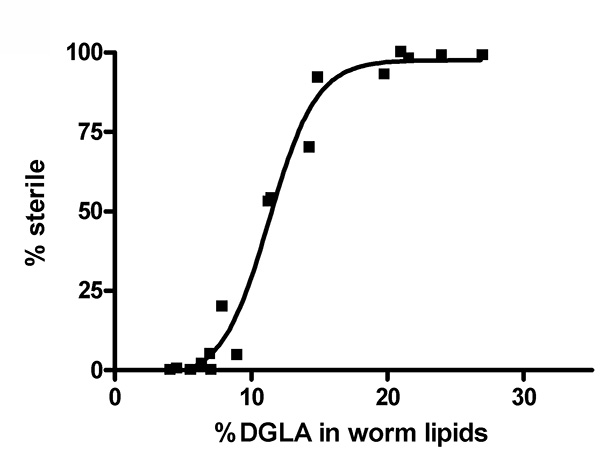
Figure 2. Increasing concentrations of DGLA in worm lipids correlate with increasing sterility in C. elegans. Wild type (N2) worms were treated with various concentrations of DGLA. The % DGLA in total worm lipids and the % of the population that is sterile is plotted for is plotted for 17 data points from five independent feeding experiments using dietary DGLA concentrations ranging from 0-0.3 mM DGLA. Click here to view larger image.
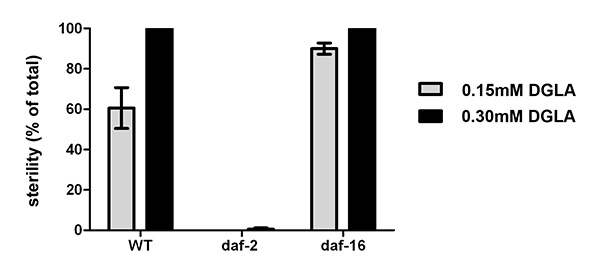
Figure 3. Physiological effects of supplementing C. elegans with DGLA. Starved L1 larval wild type, daf-2(e1370), or daf-16(mu86) were plated onto un-supplemented, 0.15 mM or 0.3 mM DGLA supplemented media and grown to the adult stage. At least 150 individual worms were then scored for sterility. Thedaf-2(e1370) mutants were almost completely fertile, even at 0.3 mM DGLA, while thedaf-16(mu86) mutants display an increased number of sterile worms compared to wild type at 0.15 mM DGLA. Error bars are SEM. Click here to view larger image.