All of the procedures involving animals were performed in compliance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals, and approved by the Institutional Animal Care and Use Committee from Georgia State University (approval number A24011). The mice were bred and maintained in the Animal Facilities at Georgia State University. The experiments were performed on a C57BL/6J strain during the perinatal period (postnatal day [P] 2-10) and included males and females. The reagents and equipment used for this study are listed in the Table of Materials.
1. Material setup
- Cut the mesh divider according to the dimensions of the cage, leaving an excess of 3 cm on the longer sides.
NOTE: The mesh provided in the table of materials is cut by the manufacturer. - Fold the edges, including the excess, in a way that creates a platform fitted precisely to the edges of the cage and with ~2.5 cm height above the floor. This will allow urine and feces to go through the mesh divider without the animals being able to retrieve the corn cob bedding. Finally, ensure that all of the sharp edges are folded down to prevent any harm to the animals.
NOTE: The mesh dividers are reusable and should be thoroughly cleaned between uses with hot water and soap, followed by spraying with 70% ethanol. - Set up the cameras on tripods and prepare the recording system.
NOTE: It is recommended to use video management software. These recordings result in .mp4 files that are automatically binned into 1 h segments. - Adjust the settings to continuous recording.
NOTE: Additional recommended settings are 1920 x 1080 resolution and 30 frames/s. - Set the resulting .mp4 files to divide into 1 h segments if not automatically done by video management software.
- If analyzing the lights-off period, use infrared lighting.
NOTE: To reduce recording glare on the side of the cage, it is recommended to disable the infrared (IR) from the camera. Instead, use IR floodlights in the room during dark phase recording.
2. Limited bedding and nesting (LBN) paradigm
- Pair 1-4 females with a single-housed male breeder.
NOTE: The recommended age for the first pairing in experimental females is P75. Using nulliparous females is ideal, but in situations where this is not feasible, such as with valuable transgenic mouse lines, multiparous females may continue breeding for approximately four months afterward. It is recommended, although not required, to check for vaginal plugs daily to confirm the day of mating (embryonic day [E]0).- Once the dams are pregnant, keep the disturbances minimal.
NOTE: Multiparous females can generally be reused to produce up to five litters before fertility declines. If the dams are reused for the experiment, they should preferentially be used for the same condition each time or only switched from Control to LBN, but never the other way around, in order to avoid any residual long-term effects of LBN on the dam.
- Once the dams are pregnant, keep the disturbances minimal.
- On E17 (or if plugs were not checked, whenever the females look evidently pregnant), separate females into their own standard plexiglass cage and give them one cotton nestlet (5 x 5 cm) to build a nest.
NOTE: It is optimal to move separated females to a quiet, small room away from the main colony room with a 12 h light/dark cycle and minimal disruptions from personnel for the Control and LBN setup. - Record the date of birth (around E19).
- At P2, count and sex the pups. Randomly assign litters to LBN or Control conditions (if it is the dam's first litter). This interaction with the pups should be completed between 1 and 4 h after the lights turn on.
- Place pups carefully into a new, clean cage to be sorted by sex. The anogenital distance between the anus and genitals is greater in males than in females and can be used as an identifier.
- Separate males and females into groups to count them.
- Cull litters with more than 8 pups and discard litters with less than 4. The optimal litter size is 4-8 pups, as anything outside of this range can interfere with the distribution of maternal care during the study.
NOTE: The variables that can be affected due to litter size are pup weight, feeding opportunity, and maternal-pup interactions, which could confound the experiment. Ideally, mice will not be cross-fostered since they are less likely to survive than rats.
- Set up the Control and LBN conditions as shown in Figure 1.
- Control: Use a standard-size mouse shoebox cage (19.4 cm x 13.0 cm x 38.1 cm) with ~220 g of corn cob bedding (depending on the cage dimensions, aiming for ~1 cm height) and one standard square cotton nestlet (5 x 5 cm).
- To help the mice acclimate to this unfamiliar environment, place one of their fecal pellets from their old cage in each corner of the new cage. In addition, place a small (dime-sized) amount of used cotton nestlet from the previous cage over the new nestlet.
- LBN: Place the previously prepared mesh divider into a standard shoebox cage with ~110 g of corn cob bedding (aiming for ~0.5 cm height). The mesh should prevent direct contact between the mice and corn cob bedding.
- Finally, add half of a cotton nestlet square (2.5 x 5 cm). Repeat all of the steps above to help with acclimation.
NOTE: It is recommended to maintain shoebox cages on standard, non-ventilated racks because it has been shown that this setup is quiet and does not induce hypothermia in pups reared in LBN cages21. However, this may not be the case with the increased airflow in ventilated cages; therefore, researchers should measure noise levels and pup core temperatures in their system if they are using ventilated cages.
- Finally, add half of a cotton nestlet square (2.5 x 5 cm). Repeat all of the steps above to help with acclimation.
- Control: Use a standard-size mouse shoebox cage (19.4 cm x 13.0 cm x 38.1 cm) with ~220 g of corn cob bedding (depending on the cage dimensions, aiming for ~1 cm height) and one standard square cotton nestlet (5 x 5 cm).
- Add the pups to their designated cage and place them on top of the nestlet. Next, place the dam in the cage facing the pups, as this will help her notice that they are present more quickly. Ensure that enough food and water is included for at least 8 days to avoid disturbing the cage until the end of the experiment at P10.
- Place a camera on a tripod in front of the cage, allowing for a clear side view of the dam and her pups. Optionally, place mirrors around the cage to better capture all of the angles. Although only 1 h of video recording per day needs to be analyzed, the camera is set up to continuously record 24/7 during the experiment so that disturbances can be monitored.
NOTE: Mice can also be observed in person, and maternal behavior can be hand-scored, although it is preferable to video-record to prevent any disruptions due to the presence of the experimenter in the room. If ventilated racks are used, it also may be necessary to use a different procedure to arrange cameras for maternal care recording than that described here. - On the morning of P10, return all of the animals to standard caging conditions (ideally identical to the control condition above) and weigh the pups.
- Treat all of the pups the same and wean at P21 into groups of 2-5 same-sex littermates.
NOTE: Aim to house same-sex littermates together whenever possible, but if necessary, house same-sex offspring from different litters in the same condition together. It is recommended that housing control and LBN offspring together be avoided (see Yang et al. for the effect of cagemate composition on behavioral phenotypes22).

Figure 1: Example of cage setup. The cage on the left side of the image shows a standard control (CTL) cage containing a full amount of bedding and a full nestlet. The cage on the right side shows a limited bedding and nesting (LBN) setup with half the amount of bedding, half a nestlet, and a mesh divider for separating the animals from the bedding. Please click here to view a larger version of this figure.
3. Maternal behavior scoring
- Although continuous recordings are collected in this setup, analyze only 1 h from P3 to P6. It is recommended to analyze videos recorded no earlier than 1 h after the lights change to allow for habituation and to analyze videos from a consistent time each day.
NOTE: The period from P3 to P6 typically contains the largest differences in maternal care due to LBN; therefore, it is only necessary to analyze these days. The suggested observation/recording time allows for the capture of activity after transitioning from the active to inactive cycle or vice versa. However, the inactive phase is when the dam is most likely to engage in maternal care and has previously been shown to contain the largest group differences in maternal behavior3.
NOTE: Capture P2 recordings later than on other days because mice need at least 1 h to habituate to the cage change. Unless this is a variable of interest, discard this day from the analysis. - Score all of the behaviors as presented in Table 1. Typically, behaviors are scored for the first 50 min after the data collection time has begun.
NOTE: This can be done by hand or electronically using the Behavioral Observation Research Interactive Software (BORIS, an open-source software), or a similar type of software. Instructions below are for hand scoring and can be adapted for whatever software is choosen. - In a printed table, record the observed behavior using the abbreviation, the start time and duration of the bout, and any descriptive notes. For example, if the dam is actively nursing and then leaves the nest but pups are still attached to her, note this as AN, followed by O. The description of the pups (and how many) still attached should be kept as a note on the side in case this is needed later.
NOTE: Any behavior under 3 s long is not analyzed. This rule helps filter out momentary behaviors due to outside disturbances, such as environmental noise. An exception is made when there is a visible interruption to an AN bout (such as by a large movement or stretch) that is quickly resumed. In this case, the behavior noted is AN, followed by N-AN, as described in Table 1. - In the event of multiple behaviors happening at once, record the most active one. An example of this is if the dam is low nursing but later begins licking and grooming, low nursing is noted to stop, and the next behavior should be marked as LG with a note that this occurred during LN.
| Type of behavior | Abbreviation | Description | ||||
| Licking / grooming | LG | The dam is engaged in licking/grooming her pups. | ||||
| Active nursing | AN | The dam is nursing her pups standing up, while her back is arched. | ||||
| New active nursing | N-AN | This behavior is used specifically when the dam interrupted nursing but quickly resumes. This is an exception to the 3s rule. | ||||
| Low nursing | LN | The dam is actively nursing her pups, but her back is low or almost flat. This behavior commonly follows AN after a period of time. | ||||
| Side Nursing | SN | The dam is lying on her side when nursing (also known as passive nursing). | ||||
| Off nest | O | The dam is not on the nest, and she is not eating/drinking. This can be observed in her walking around the cage or exploring. | ||||
| Eating/drinking | E | The dam is off the nest eating or drinking. | ||||
| Self-grooming | SG | The dam is grooming herself. | ||||
| Carrying pups | C | The dam carries the pups, usually to relocate them back to the nest. | ||||
| Nest building | NB | The dam is actively constructing or relocating the nest. | ||||
| Move on nest | M | The dam is moving on the nest. This presents with the dam interacting with the pups in a way different than LG or any type of nursing, such as sniffing, rearing, or stepping on the pups. | ||||
Table 1: Description of the maternal care behaviors.
4. Maternal behavior data analysis
- Compile the scoring results into a spreadsheet.
- If the videos were analyzed electronically, delete the behaviors with bouts shorter than 3 s, as described in step 3.
- Calculate the average bout length, average frequency, and total duration of licking and grooming for each day of observation.
NOTE: Descriptive statistics can be performed on any of the scored behaviors, but licking and grooming will typically present the most evident disruption in terms of fragmentation by LBN (i.e., shorter, more frequent bouts). These variables can be analyzed as an average across P3-P6, or as repeated measures by day to look for any changes over time if this is of interest.
5. Calculation of entropy
NOTE: Entropy, or unpredictability, of maternal care behaviors is calculated based on the method proposed by Vegetabile et al.23. This method is based on the assumption that maternal care behaviors act as a Markov chain, which can be used to estimate the entropy rate of a behavioral sequence. Each dam's sequence of behaviors is characterized using the empirical transition matrix <pij> i,j = 1…7 of conditional probabilities of moving from one behavior (i) to another behavior (j), and the entropy rate is calculated from this as previously described3,23 and as follows:
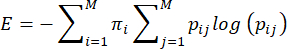
where pij is the conditional probability that behavior j is observed next after a dam is observed performing behavior i, πi is the frequency with which behavior i is observed, and M (=7) is the total number of different behaviors. The reader is referred to Vegetabile et al.23 for a discussion of the theoretical underpinnings of the equations; here, the focus is on how to apply the method in the LBN model.
- To compute this, arrange the format required for the analysis.
- Combine AN, LN, SN, and N-AN into a single variable named N by adding the times together, as they all involve nursing.
NOTE: Refer to Table 1 for the description of the maternal care behaviors. - Add the behaviors that are not related to self- or pup-directed behaviors (M and O) into O.
- Input LG, E, SG, C, and NB separately to result in 7 total behaviors.
- Combine AN, LN, SN, and N-AN into a single variable named N by adding the times together, as they all involve nursing.
- Create a spreadsheet with columns following this order: Mouse ID, Litter ID, Litter #, Treatment, Day, Time, Behavior, and Status.
- In the mouse ID, add the mouse line/genotype of the dam.
- In litter ID, use the dam's identifier followed by the litter number.
- In litter #, indicate the number of litters the dam has had.
- Ensure that the treatment is LBN or Control, according to the conditions in which the animals were placed.
- Indicate the day as the corresponding postnatal day analyzed.
- Indicate the time as the time stamp in the video when the behavior started and ended (relative to the start of recording).
- Select the behavior from the seven described earlier (N, O, LG, E, SG, C, and NB) (Table 1).
- Mark the two types of statuses, START and STOP. Therefore, each behavior will appear twice; the first notation marks the start, and the second marks the stop.
- Following this format, include the information of all the analyzed days.
- In the R environment, import the datasets that have been formatted as above.
- Install the packages available at https://github.com/bvegetabile/entropyRate.
NOTE: Running this code results in a folder named LBN; regardless of the condition, this folder contains the calculated entropy. Additionally, the entropy of each day can then be averaged by the subject for P3-P6 and compared between conditions.
The representative results demonstrate how ELA, imposed by an impoverished environment in LBN cages, affects maternal care from dams and offspring physiological outcomes. The daily entropy in maternal care behavior is higher in LBN across days P3-P6 (F1,58 = 7.21, p = 0.0094; Figure 2A), as well as the average entropy of each dam from this time period (t15 = 3.03, p = 0.0085; Figure 2B). Notably, there is no significant difference in average entropy rate across different litters from the same dam when maintained within the same treatment group (F1.699,4.247 = 0.57, p = 0.58; Figure 2C), suggesting that entropy rate may be a somewhat stable trait for each dam. Amongst all of the behaviors, licking and grooming is the one that has been shown to be most fragmented by LBN3. The LBN dams display a higher frequency of licking and grooming their pups (LG) (t16 = 4.04, p = 0.0010; Figure 2D) and in shorter bouts (t16 = 3.25, p = 0.0050; Figure 2E). However, there is no significant difference in the total duration of LG between the Control and LBN dams (t16 = 1.52, p = 0.15; Figure 2F).
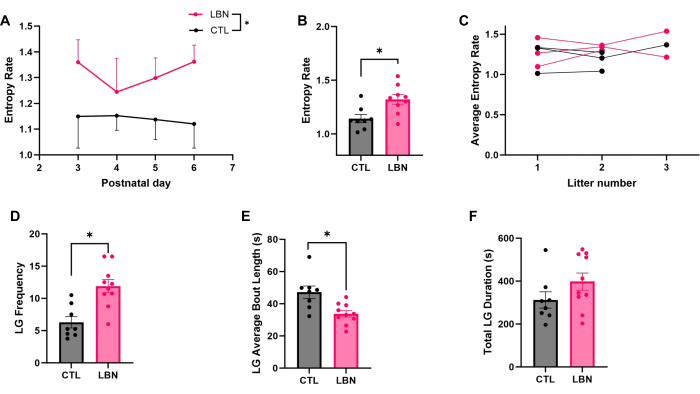
Figure 2: Maternal behavior analysis. (A) The daily entropy rate is higher in limited bedding and nesting (LBN) dams vs. control (CTL) as analyzed by the mixed-effects model (F1,58 = 7.21, p = 0.0094). (B) The average entropy rate is higher in LBN dams vs. CTL (t15 = 3.03, p = 0.0085). (C) The average entropy rate (P3-P6) is not significantly different across multiple litters within the same dam when maintained in the same treatment group (F1.699,4.247 = 0.57, p = 0.58). Each line represents a single dam. (D) The cumulative frequency of licking and grooming (LG) events is higher for LG dams vs. CTL (t16 = 4.04, p = 0.0010). (E) The LG average bout length is shorter for LG dams vs. CTL (t16 = 3.25, p = 0.0050). (F) The cumulative time spent on LG is not significantly different due to LBN (t16 = 1.52, p = 0.15). Data are mean ± SEM, * p < 0.05. Please click here to view a larger version of this figure.
Pups that were reared in LBN conditions are significantly smaller at P10 (t61 = 6.30, p < 0.0001; Figure 3A). This difference typically persists at weaning age (t62 = 6.29, p = <0.0001; Figure 3B) but is no longer observed by adulthood (t38 = 1.08, p = 0.29; Figure 3C). However, corticosterone levels are increased at baseline in adulthood (t18.79 = 2.23, p = 0.038; Figure 3D), suggesting enduring physiological effects of LBN. These are the expected differences in a successful experimental setup. In the case of a suboptimal setup, the values of LG bout length, entropy, and pup weights may present no differences, likely due to a stressed "Control" group. For the purposes of this paper, sex was collapsed when analyzing offspring outcomes because sex differences are not typically observed in the physiological outcomes described here; however, sex differences in other types of outcomes, such as cognitive and emotional behavior, are commonly reported in this model and should be investigated further4.
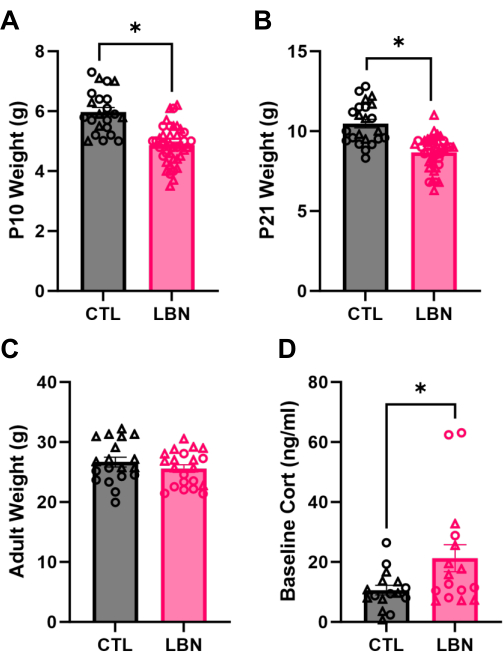
Figure 3: Offspring outcomes. (A) Limited bedding and nesting (LBN) decreases pup weight measured at P10, just before return to standard cages (t61 = 6.30, p < 0.0001). (B) Weight is still decreased by LBN at the weaning age (t62 = 6.29, p < 0.0001). (C) Adult weight no longer differs due to LBN (t38 = 1.08, p = 0.29). (D) The baseline concentration of corticosterone in adulthood is increased by LBN, as analyzed by Welch's t-test (t18.79 = 2.23, p = 0.038). For all graphs, females are shown with circles and males with triangles. Data are mean ± SEM, *p < 0.05. Please click here to view a larger version of this figure.
