The presented flowchart illustrates the stable condition of the mustard aphids from field collection to virulence screening. The maintenance of aphids from field collection ensured a stable increase in aphid colonies with an adequate food supply. The field-collected aphids were confirmed as mustard aphids through the use of molecular markers, including PCR amplicon size and LeCO1 sequencing. The virulence screening, conducted using the detached-leaf method, revealed a consistent survival rate for mustard aphids, with the control group exhibiting an 85% survival rate (Figure 4).
During the virulence screening, Cc-NCHU-213 demonstrated the fastest aphid-killing ability, resulting in 50% and 90% mortalities at 3 days and 4.5 days post-inoculation (d.p.i.), respectively (Figure 4). However, varying aphid-killing abilities were observed among the five B. bassiana isolates. Bb-NCHU-141, -143, and -153 exhibited slow aphid-killing abilities, with only 5% mortality at 3 d.p.i., even when excluding the effects of 0.03% Tween 80 spraying or other lethal factors (Figure 4). Hence, the corrected mortality formula was employed to normalize the control mortality. Most EPF isolates against mustard aphids exhibited corrected mortality rates higher than 70% within 5 d.p.i., except for Pl-NCHU-152 (Table 3). Among these EPF isolates, Bb-NCHU-286 demonstrated the highest mortality rate of 100% (Table 3). Additionally, EPF mycosis was observed on cadavers of mustard aphids infected with Metarhizium spp., Beauveria spp., Purpureocillium lilacinum, and Cordyceps cateniannulata during the virulence screening, indicating the effectiveness of this system (Figure 5).
Based on the results of the virulence screening, two EPF isolates, namely Mb-NCHU-197 and Cc-NCHU-213, exhibiting rapid insect-killing activity (40% and 50% mortality at 3 d.p.i., respectively), were selected for bioassay against mustard aphids. The results demonstrated significantly different corrected mortalities for Mb-NCHU-197 and Cc-NCHU-213 at 3 and 4 d.p.i. with an inoculation of 107 conidia/mL (Figure 6). In the LT50 assay, the treatment with 107 conidia/mL of Cc-NCHU-213 exhibited a significantly shorter duration compared to other treatments (Table 4). Furthermore, the LC50 value of Cc-NCHU-213 (9.32 × 104) was lower than that of Mb-NCHU-213 (2.30 × 105), indicating that Cc-NCHU-213 possesses greater virulence against mustard aphids (Table 5).
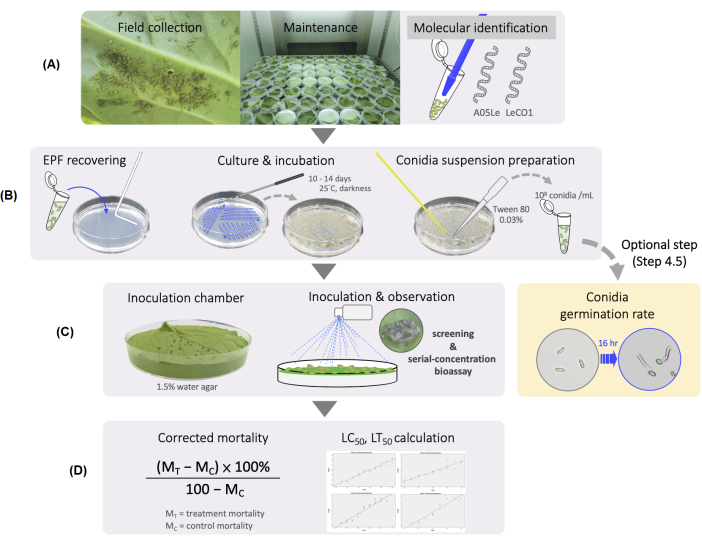
Figure 1: Experimental flowchart for screening EPF virulence against mustard aphids. (A) Establishment of a mustard aphid-rearing system. (B) Preparation of EPF. (C) Fungal inoculation. (D) Statistical analysis. Please click here to view a larger version of this figure.

Figure 2: Electrophoresis of mustard aphid genomic DNA amplified with A05Le and Le CO1 primer sets. Electrophoresis was performed on a 1% agarose gel. M = 100 bp DNA ladder; bp = base pairs.The red asterisk indicates the target bend of PCR amplification. Please click here to view a larger version of this figure.

Figure 3: Differences between apterous fourth-instar nymph and adult mustard aphids. (A) Fourth-instar nymph. (B) Adult. The tibiae of the hind legs of the fourth-instar nymph are whitish (marked with red arrow). Newly emerged aphids were removed by fine camel brush during virulence tests. Scale bar = 1 mm. Please click here to view a larger version of this figure.
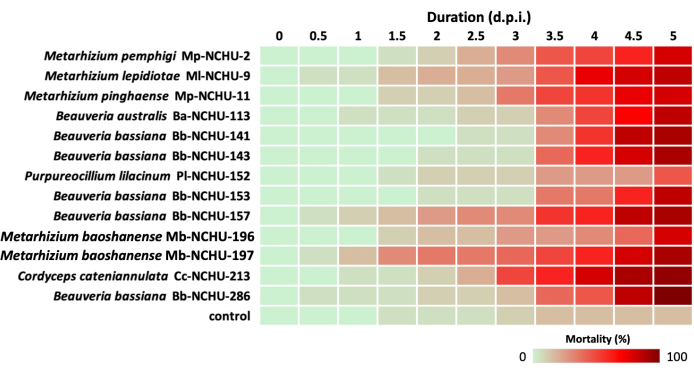
Figure 4: Mortality heat map of 13 EPF isolates against mustard aphids through the detached-leaf method. Please click here to view a larger version of this figure.

Figure 5: Observation of fungal mycosis of 13 EPF isolates. Mepe-NCHU-2 = Metarhizium pemphigi; Mp-NCHU-11 = Metarhizium pinghaense; Mb = Metarhizium baoshanense;Cc = Cordyceps cateniannulata; Ba = Beauveria australis;Bb = Beauveriabassiana; Pl = Purpureocillium lilacinum. Scale bar = 1 mm. Please click here to view a larger version of this figure.
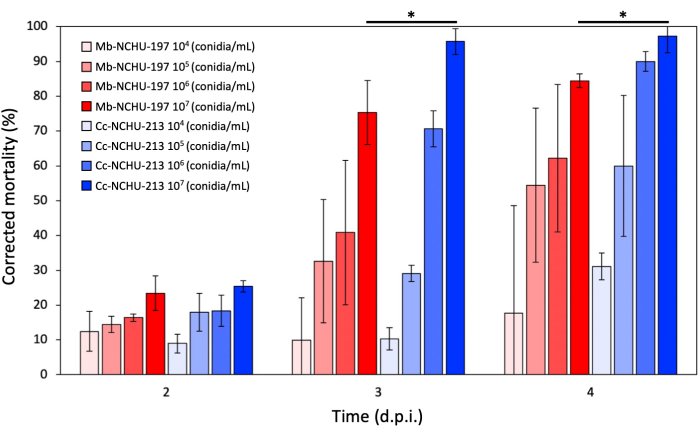
Figure 6: Corrected mortality of fungal isolates Mb-NCHU-197 and Cc-NCHU-213 against mustard aphid. The error bars represent the standard deviation (SD). The mortalities of Mb-NCHU-197 and Cc-NCHU-213 at the same time point with the same inoculated concentration were compared using independent t-test, and mortalities marked with an asterisk were found to be significantly different (p < 0.05). Please click here to view a larger version of this figure.
| Isolate | Species | Host or source* | Location |
| Mp-NCHU-2 | Metarhizium pemphigi | soil | Yilan |
| Ml-NCHU-9 | Metarhizium lepidiotae | soil | Yilan |
| Mp-NCHU-11 | Metarhizium pinghaense | soil | Yilan |
| Ba-NCHU-113 | Beauveria australis | soil | Taichung |
| Bb-NCHU-141 | Beauveria bassiana | Hypothenemus hampei | Chiayi |
| Bb-NCHU-143 | Beauveria bassiana | Hypothenemus hampei | Chiayi |
| Pl-NCHU-152 | Purpureocillium lilacinum | Tessaratoma papillosa | Chiayi |
| Bb-NCHU-153 | Beauveria bassiana | Rhynchophorus ferrugineus | Chunghua |
| Bb-NCHU-157 | Beauveria bassiana | Rhynchophorus ferrugineus | Chunghua |
| Mb-NCHU-196 | Metarhizium baoshanense | soil | Taichung |
| Mb-NCHU-197 | Metarhizium baoshanense | soil | Taichung |
| Cc-NCHU-213 | Cordyceps cateniannulata | soil | Taichung |
| Bb-NCHU-286 | Beauveria bassiana | Cerambycidae | Taichung |
Table 1: EPF isolates used in this study.
| Primer name | Sequence (5' – 3') | Product size (bp) | Reference | |
| A05Le | F-GGGTCTTGGATGGTGTGGTG | 953 | Lu et al. [21] | |
| R-AGGGGTCTTGTCGCCATTTT | ||||
| LeCO1 | F-CTTTTCCCATGATCAATTTT | 593 | This study | |
| R-ACGTAGTGGAAATGAGCAAC | ||||
Table 2: Primer pairs used for molecular identification of mustard aphids.
| Isolate | Species | Corrected mortality (%) |
| Mp-NCHU-2 | Metarhizium pemphigi | 76.47 |
| Ml-NCHU-9 | Metarhizium lepidiotae | 82.35 |
| Mp-NCHU-11 | Metarhizium pinghaense | 76.47 |
| Ba-NCHU-113 | Beauveria australis | 82.35 |
| Bb-NCHU-141 | Beauveria bassiana | 88.24 |
| Bb-NCHU-143 | Beauveria bassiana | 88.24 |
| Pl-NCHU-152 | Purpureocillium lilacinum | 35.29 |
| Bb-NCHU-153 | Beauveria bassiana | 82.35 |
| Bb-NCHU-157 | Beauveria bassiana | 88.24 |
| Mb-NCHU-196 | Metarhizium baoshanense | 76.47 |
| Mb-NCHU-197 | Metarhizium baoshanense | 88.24 |
| Cc-NCHU-213 | Cordyceps cateniannulata | 94.12 |
| Bb-NCHU-286 | Beauveria bassiana | 100.00 |
Table 3: Corrected mortality rates of 13 EPF isolates against mustard aphids at 5 days post inoculation (d.p.i.).
| Isolate | Species | Conc.(conidia/mL) | N* | LT50 (days)† | 95% confidence limits | Slope (SE) | X2 (df)‡ |
| Mb-NCHU-197 | Metarhizium baoshanense | 104 | 60 | 3.816 a | 3.61–4.05 | 0.60 (0.05) | 15.64 (28) |
| 105 | 60 | 3.112 bd | 2.95–3.28 | 0.72 (0.05) | 14.61 (28) | ||
| 106 | 60 | 2.908 b | 2.76–3.06 | 0.85 (0.06) | 15.04 (28) | ||
| 107 | 60 | 2.549 c | 2.40–2.69 | 0.90 (0.06) | 24.31 (28) | ||
| Cc-NCHU-213 | Cordyceps cateniannulata | 104 | 60 | 3.948 a | 3.71–4.23 | 0.53 (0.05) | 8.81 (28) |
| 105 | 60 | 3.237 d | 3.07–3.41 | 0.72 (0.05) | 22.86 (28) | ||
| 106 | 60 | 2.414 c | 2.28–2.54 | 1.08 (0.07) | 28.30 (28) | ||
| 107 | 60 | 2.132 e | 2.02–2.25 | 1.41 (0.10) | 28.96 (28) | ||
| *Number of insects observed. | |||||||
| †LT50 values marked with different letters were considered significantly different as their 95% confidence limits did not overlap. | |||||||
| ‡X2, Chi-square value in Pearson's goodness-of-fit test; df, degrees of freedom | |||||||
Table 4: LT50 values of fungal isolates Mb-NCHU-197 and Cc-NCHU-213 against mustard aphids under different conidia concentrations. *Number of insects observed. †LT50 values marked with different letters were considered significantly different as their 95% confidence limits did not overlap. ‡X2, Chi-square value in Pearson's goodness-of-fit test; df, degrees of freedom.
| Isolate | Species | N* | LC50 (conidia/mL)† | 95% confidence limits | Slpoe (SE) | X2 (df)‡ |
| Mb-NCHU-197 | Metarhizium baoshanense | 240 | 2.30 × 105 a | 8.63 × 104–5.70 × 105 | 0.43 (0.08) | 10.14 (10) |
| Cc-NCHU-213 | Cordyceps cateniannulata | 240 | 9.32 × 104 a | 4.97 × 104–1.62 × 105 | 0.76 (0.09) | 4.33 (10) |
| *Number of insects observed. | ||||||
| †LC50 values marked with different letters were considered significantly different as their 95% confidence limits did not overlap. | ||||||
| ‡X2, Chi-square value in Pearson's goodness-of-fit test; df, degrees of freedom | ||||||
Table 5: LC50 values of fungal isolates Mb-NCHU-197, and Cc-NCHU-213 against mustard aphids. *Number of insects observed. †LC50 values marked with different letters were considered significantly different as their 95% confidence limits did not overlap. ‡X2, Chi-square value in Pearson's goodness-of-fit test; df, degrees of freedom.
