The process of generating lung tissue slices involves several key steps, including preparation, embedding, and sectioning. The agarose solution (3% (w/v) is made by dissolving 1.5 g of low-melting-point agarose in 50 mL of sterile PBS. Consumables needed include a plastic cup, lid from a 50 mL conical tube (rim removed), forceps, and scissors. The lung tissue is carefully resected into smaller blocks with fine forceps and sterile scissors (Figure 1A,B). Next, to embed the lung tissue blocks, the bottom of the cup is filled with a small amount of the agarose, a lid is placed on the surface of the agarose, and the cup is stored at 4 °C for 5 min (Figure 1C). Approximately 1 mL of liquid agarose is added to the lid, and the lung tissue block is placed onto the lid. The agarose is left for 2-3 min to semi-solidify at room temperature (Figure 1D). Once a lung block is held in place by the semi-solidified agarose, liquid agarose is then poured into the cup until the lung tissue block is fully submerged. This is stored at 4 °C for 15 min (Figure 1E,F). The low-melting-point agarose embedding provides the necessary support and stiffness to the lung tissue, which helps maintain its original architecture during the slicing process. A sharp blade is used to remove excess agarose surrounding the lung tissue. A uniform layer of approximately 5 mm thickness is left around the tissue (Figure 1G). The excised lung tissue block is then carefully glued onto the tissue holder (Figure 1H). The vibratome buffer tray is filled with PBS, ice is placed in the surrounding ice bath, and the specimen disk installed in the buffer tray (Figure 1I). The vibratome cutting parameters set on the panel were: slice thickness: 200 µm; frequency: 100 Hz; amplitude of the blade: 1.3mm; forward speed of the blade: 0.02-0.03 mm/s, which depends on tissue stiffness (Figure 1J). Finally, staining is carried out on free-floating slices (Figure 1K–M).
Immunofluorescence staining was carried out in 200 µm sections from each species for smooth muscle actin (SMA, a smooth muscle cell and myofibroblast marker), elastin (extracellular matrix protein) and Lycopersicon Esculentum lectin (LEA), which binds to bronchoalveolar epithelial cells. The lung is a structurally heterogeneous tissue and as such, differential SMA and elastin distribution is observed across structures, including alveolar ducts, blood vessels, intrapulmonary bronchioles, and alveoli from mice, pigs, and humans (Figure 2A-L). A high-resolution rendering of a 20 µm volume of a bronchiole shows how fine structures within the lung can be examined at a semi-three-dimensional level (Supplemental Video S1). As a quantitative example, utilizing image tracing, we show how the size of the alveoli differs between samples (n = 20 alveoli) (Figure 2L,M).
To show how it is possible to quantitatively utilize these advanced imaging data, we compared type 2 pneumocyte (TII) cell distributions in human lung tissue from an adult and an infant. The human-specific type 2 alveolar cell antibody, HTII-280, has a strong affinity to the surface of human type 2 cells (Figure 3A-F). These cells can further be viewed in the three-dimensional space of a single alveolus using volumetric imaging (Supplemental Video S2). To quantify the TII number between the adult and infant samples, we processed the HTII-280 count across 10 random fields of view (fov). Between these samples, the infant sample yielded a significantly higher TII cell count (Figure 3G). However, the infant sample also exhibited a significantly higher scaffold coverage than the adult, which could explain the higher cell number (Figure 3H). To account for this, we then assessed the number of TIIs based on scaffold coverage, as cells per mm2 of tissue, which still maintained a significantly higher number of TIIs in the infant sample (Figure 3I). Thus, the higher TII count in the infant sample is not due to increased scaffold coverage and highlights the importance of assessing cell distributions in the lung based on the area of the scaffold, not the overall fov.

Figure 1: Protocol for cutting lung tissue slices. (A) The solutions and materials: a 3% (w/v) agarose solution obtained by dissolving 1.5 g of low-melting-point agarose in 50 mL of sterile phosphate-buffered saline; a plastic cup; a lid from 50 mL conical tube; forceps, and scissors. (B) The lung tissues were dissected by using fine forceps and sterile scissors. (C) Fill the bottom of the cup with a small amount of agarose and place the lid on the surface of the agarose; then, store it at 4 °C for 5 min. (D) Add 1-2 mL of liquid agarose to the lid slowly, place the lung tissue chunk onto the lid carefully, and store it at 4 °C for 2 min. (E,F) Pour the liquid agarose into the cup until the lung tissue chunk is fully submerged, store it at 4 °C for 15 min, and then break the plastic cup gently. (G) Use a sharp blade to remove excess agarose surrounding the lung tissue, leaving a uniform layer of approximately 5 mm thickness around the tissue. (H) The excised lung tissue block is then carefully glued onto the tissue holder. (I) Fill the vibratome buffer tray with PBS, place ice in the surrounding ice bath, and install the specimen disk into the buffer tray. (J) Set cutting parameters on the panel of the vibratome; slice thickness: 200 μm, frequency: 100 Hz, amplitude of the knife: 1.3 mm, and forward speed of the blade of 0.02-0.04 mm/s. (K–M) Representative images of a free-floating slice still embedded in agarose and ready for immunofluorescence staining. Please click here to view a larger version of this figure.
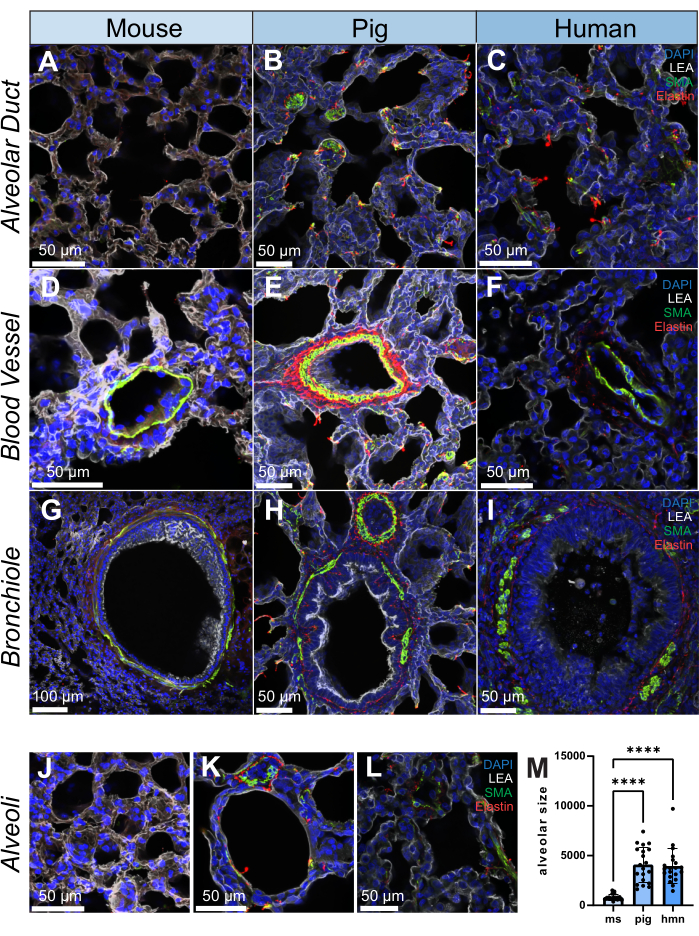
Figure 2: Immunofluorescence images of lung structure in human, pig, and mouse. (A-L) Representative images of an alveolar duct, blood vessel, bronchiole, and alveoli in lung tissue slices from mouse, pig, and human, respectively. SMA (shown in green) is a marker for smooth muscle cells and is observed in the vascular and bronchial walls of lung tissue. Elastin (shown in red) forms part of the extracellular matrix of the lungs. (M) Quantification of alveolar size by selecting 20 random positions on each lung tissue slice for both human, pig, and mouse samples. Histograms were utilized to illustrate the distribution of alveolar size within each group. Scale bars = 50 µm (A–F, H–L), 100 µm (G). Kruskall-Wallis Test. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Abbreviations: DAPI = 4′,6-diamidino-2-phenylindole; LEA = Lycopersicon Esculentum lectin; SMA = smooth muscle actin; ms = mouse; hmn = human. Please click here to view a larger version of this figure.
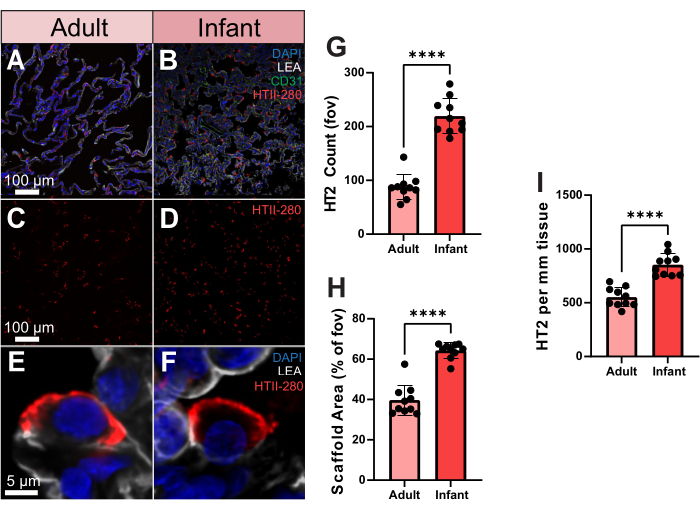
Figure 3. Type 2 pneumocyte distribution in human adult and infant samples. (A,B) Representative images of adult and infant human lung tissue stained for HTII-280 (shown in red), CD31 (shown in green), and LEA (shown in gray). (C,D) Representative images of adult and infant human showing type 2 pneumocyte distribution alone. (E,F) Single-cell resolution representative images of an individual type 2 pneumocyte from adult and infant human lung samples. (G) Quantification of the number of type 2 pneumocytes between adult and infant samples. (H) Quantification of lung tissue scaffold area coverage (% vs air) in adult and infant samples. (I) Quantification of type 2 pneumocyte per mm2 of tissue in adult and infant samples. Scale bars = 5 µm (E,F), 100 µm (A–D). n = 10 fov per sample. Unpaired t-test or Mann-Whitney Test. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Abbreviations: DAPI = 4', 6-diamidino-2-phenylindole; LEA = Lycopersicon Esculentum lectin; HTII = human type II cells; CD31 = Platelet endothelial cell adhesion molecule-1/cluster of differentiation 31; fov = field of view. Please click here to view a larger version of this figure.
Supplemental Video S1: Video of 20 µm imaging volume of a single porcine airway stained using DAPI, LEA, SMA, and elastin showing a high-resolution view of the structure of airway wall elements. Abbreviations: DAPI = 4′,6-diamidino-2-phenylindole; LEA = Lycopersicon Esculentum lectin; SMA = smooth muscle actin. Please click here to download this File.
Supplemental Video S2: Video of 30 µm imaging volume of a single alveolus stained using DAPI, LEA, and HTII-280 showing type 2 pneumocyte distribution across the structure. Abbreviations: DAPI = 4′,6-diamidino-2-phenylindole; LEA = Lycopersicon Esculentum lectin; SMA = smooth muscle actin. Please click here to download this File.
