All procedures were approved by the Committee for Ethics in Experiments with animals of the University of Antioquia (UdeA) (minutes 104 of June 21st, 2016, and 005 of April 15th, 2021), according to Law 84 of 1989 and Resolution 8430 of 1993 issued by the Colombian Government and were performed and reported in compliance with the Animal Research: Reporting In Vivo Experiments (ARRIVE) guidelines41. All results presented here come from healthy, 7-13 weeks old, 20-26 g, C57BL/6 male mice. Figure 1 shows the general design of this study and the order of the procedures. All reagents, materials, and equipment details are listed in the Table of Materials.
1. Animals
- House a maximum of six mice per acrylic, transparent, rectangular cage, with wood-derived bedding, under conditions of controlled temperature (21 ± 2 °C) and light:darkness (12:12 h) cycles.
- Give the animals free access to food and tap water in specific pathogen-free animal facilities with no environmental enrichment.
2. Dissection
- Solutions, materials, and reagents
- Prepare and filter (0.22 µm) the working solutions with the following composition (all concentrations in mM):
- Tyrode: 5.4 KCl, 1 MgCl2, 140 NaCl, 0.33 NaH2PO4, 2 CaCl2, 10 glucose, 10 HEPES, pH 7.3
- Dissociation: 2.7 KCl, 1.2 KH2PO4, 0.5 MgCl2, 138 NaCl, 0.1 Na2HPO4, 1 CaCl2, pH 7.4
- Phosphate-buffered saline (PBS): 137 NaCl, 8.6 Na2HPO4, 2.8 KH2PO4, pH 7.34
- Prepare two dissection chambers; stereoscope; operating scissors; fine scissors; fine forceps; and clean, transparent, non-conic, 1-1.5 cm wide, glass vials of 3-4 mL total volume with caps. Arrange a system for electrically stimulating the muscles in the dissection chambers.
- Prepare fire-polished Pasteur glass pipettes of different width tips: 5, 4, 3, 2, and 1 mm.
- Set the water bath to 37 °C. Weigh aliquots of 3 mg of collagenase type 2.
- Prepare and filter (0.22 µm) the working solutions with the following composition (all concentrations in mM):
- Procedure
- Sacrifice the mouse using methods approved by the local Ethics Committee. Cervical dislocation is recommended because it is rapid, less stressful, and avoids exposure to drugs, which may affect the muscle tissue (such as CO2 or some anesthetics). Start dissection immediately to obtain better results.
- Place the mouse on a foam surface and tape or pin the forelimbs. Cut both hindlimbs over the knees with the operating scissors, transfer each of them to a separate dissection chamber, and add cold (10-20 °C) Tyrode to cover the tissue.
NOTE: Each hindlimb will give six different muscles in the following order: FDB, soleus, EDL, EHL, PL, and PDQA. Detailed anatomical references to dissect the six muscles intact from tendon to tendon are given in Figure 2 and elsewhere42. - Pin the first hindlimb to the dissection chamber in a position in which the posterior face of the legs is visible. Remove the skin under magnification; then expose and remove the FDB (Figure 2). Store it in one labeled glass vial with 1 mL of Tyrode solution.
NOTE: Appropriate magnification and previous training are required to avoid any undesired cut in the muscle tissue. - Expose, remove, and store the soleus in a separate vial with 1 mL of Tyrode. Use fine scissors to first separate the gastrocnemius and then to remove the soleus, as indicated in Figure 2.
- Expose the anterior face of the leg, remove the skin, and identify the distal tendons of the tibialis anterior and the EDL muscles in the ankle. Remove and discard the tibialis; then cut the distal tendons of the EDL (Figure 2). Continue dissection until removing the EDL and place it in a separate glass vial with 1 mL of Tyrode.
- Remove the EHL muscle, which lies just posterior and medial to the EDL. Start dissection by identifying and following the tendon to the 1st digit, as indicated in the corresponding panel of Figure 2. Keep the muscle in a separate glass vial with 1 mL of Tyrode.
- Identify and follow the most external tendon of the peronei to cut it and remove the PL muscle (Figure 2). Place the muscle in a separate glass vial with 1 mL of Tyrode.
- Identify and follow the tendon to the 4th digit; cut it and remove the PDQA muscle (Figure 2). Place it in a separate glass vial with 1 mL of Tyrode.
- Repeat the procedure with the second hindlimb.
- Gather both muscles of the same type in one labeled glass vial or a small Petri dish with Tyrode solution.
NOTE: If more than two pairs of muscles are planned to be dissected during a working session, recruit two researchers for the dissection procedure.
3. Muscle fiber isolation protocol
- Renew the Tyrode solution in the dissection chambers to remove debris and mouse fur. Pour the FDB muscles into one dissection chamber, verify their integrity, and transfer them to a new glass vial with 1 mL of dissociation solution. Repeat this procedure with the EHL, PL, and PDQA muscles.
NOTE: If a muscle looks hypercontracted, cut, or is unresponsive to the electrical stimulation, do not continue to the next protocol step. Instead, optimize the dissection protocol by verifying the quality of the solutions (pH, contamination, osmolarity) and gaining more dissection skills (Figure 1C and Supplemental Video S1). - Perform longitudinal or diagonal cuts to the soleus and EDL muscles, following the orientation of the fibers (Figure 2). For the soleus, follow the central tendon, cutting ~80% of its length. For EDL, just follow one or two tendons and cut about the same length as for soleus. Put each pair of muscles in glass vials with 1 mL of dissociation solution.
NOTE: This procedure makes the EDL and soleus smaller and allows for the collagenase to better enter the tissue. Sufficient magnification (40-50x), as well as fine scissors and forceps, are mandatory. Always check sample integrity by visual inspection and electrical stimulation before continuing to the next step of the dissociation protocol. - Add 3 mg of collagenase type 2 (with an activity of 250-300 U/mg) to each vial containing 1 mL of dissociation solution and a pair of muscles. Standardize the exact amount of collagenase by considering the activity of the enzyme batch used.
- Incubate the pairs of muscles in the water bath for 65-90 min at 36.8-37 °C, with gentle shaking.
NOTE: Be rigorous with the temperature control. Standardize the procedure so that the muscles do not remain in collagenase for more than 100 min to avoid damage. - Check the vials under stereoscope magnification every 5 min after the 65th min of incubation. When the muscles look slightly rippled, ragged, and loose, gently shake the vial and verify if some fibers start detaching readily. If this is the case, wash the muscles with Tyrode at room temperature to inactivate and remove the collagenase.
NOTE: Washing must be done carefully, without touching the muscles with the pipettes. Start by adding 0.8 mL of Tyrode and then remove 0.8 mL of the solution. Repeat this procedure 4-5x and verify that the solution becomes fully transparent. - Separate more fibers from the bulk of the muscles with very gentle trituration in Tyrode with the help of the set of fire-polished Pasteur pipettes. Start by agitating the solution around the muscle with the widest pipette (5 mm tip) and then gently pull the muscles up into and out of the pipette 3-4x. When the muscle starts releasing fibers and becomes thinner, repeat the procedure with the next pipette (4 mm tip).
NOTE: Fibers rendered via this procedure remain excitable and contract briskly for more than 24 h, as exemplified using PL, EDL, EHL, and soleus fibers in Supplemental Video S2, Supplemental Video S3, Supplemental Video S4, and Supplemental Video S5.
4. Experimental procedures
NOTE: Isolated fibers were used for sarcoplasmic Ca2+ concentration estimations, morphometric measurements, and myosin heavy chain (MHC) expression studies.
- Measurement of the sarcoplasmic Ca2+ concentration during a twitch
- Mount a clean, glass slide on the experimental bath chamber. Coat the slide with 2-3 µL of laminin and allow it to dry for 30 s before pouring ~400 µL of the fiber suspension onto the slide. Allow the fibers to adhere to the laminin for 10-15 min at room temperature.
- Mount the experimental chamber onto the stage of an inverted microscope equipped for epifluorescence (Figure 3A).
- Evoke single twitches to verify the viability of the fibers by applying rectangular current pulses (0.8-1.2 ms) through the two platinum electrodes placed along either side of the experimental chamber. Even when attached to laminin, the contraction of the fibers is still visible mainly at the extremes.
- Load the fibers with 3.5-4.5 µM of the fast Ca2+ dye Mag-Fluo-4, AM for 4-5 min in Tyrode solution. After this time, gently wash with Tyrode to remove the extracellular dye. Allow the intracellular dye to be de-esterified for ~15-20 min under dark conditions. Always keep the temperature below 22 °C to avoid the dye compartmentalization.
NOTE: Prepare a stock of Mag-Fluo-4, AM in dimethyl sulfoxide (DMSO) only, so that the final concentration of DMSO in the loading Tyrode solution is less than 0.5%. - Illuminate the fiber with a white light-emitting diode (LED) and a filter set with the following wavelengths for excitation/dichroic/emission: 450-490/510/515 nm (Figure 3A).
NOTE: Alternative sources of excitation include mercury and xenon fluorescence lamps. Use the lowest possible intensity and size of the excitation spot to avoid photobleaching of the dye and damage to the cell. - Evoke the fiber´s Ca2+ response (sarcoplasmic Ca2+ transients) by applying rectangular current pulses (0.8-1.2 ms) through the two platinum electrodes placed along either side of the experimental chamber at 20-22 °C.
- Collect and save the light signals with an oil immersion 40x long-distance objective suitable for fluorescence and a photomultiplier tube (PMT) connected to a digitizer (Figure 3A and Supplemental Video S6). Ensure a scale in the acquisition software of 0-200 arbitrary units (AU) and set the resting fluorescence (Frest) of the experiment to 10 AU on that scale by modulating the size of the excitation spot and the gain of the PMT. Once the procedure is standardized, keep the gain unmodified from one experiment to the other and set the scale only through minor adjustments in the spot size.
NOTE: If movement artifacts arise, use 20-30 µM N-benzyl-p-toluene sulphonamide (BTS) in the Tyrode solution. - Analyze and calibrate the signals as follows:
- Lowpass filter the whole trace at 1 kHz.
- Calculate the Frest in 1 s of the trace, adjust the Frest to 0, and measure the peak sarcoplasmic Ca2+ transients ' amplitude (Fpeak). Present the amplitude as in equation (1):
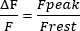 (1)
(1) - Calculate the peak Ca2+ concentration ([Ca2+], µM) using equation (2)26 and the following parameters: in situ dissociation constant (Kd) = 1.65 × 105 µM2, maximum fluorescence (Fmax) of 150.9 AU, minimum fluorescence (Fmin) of 0.14 AU, Mag-Fluo-4 concentration [D]T of 229.1 µM26. Fpeak was already obtained in step 4.1.8.2.
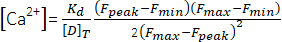 (2)
(2) - Measure the rise time from 10% to 90% of the amplitude (RT, ms), the duration at half maximum (HW, ms), and the decay time from 90% to 10% of the amplitude (DT, ms). Then, estimate the decay kinetics according to a fit with the biexponential function (equation 3):
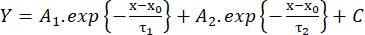 (3)
(3) - Save the values of the time constants of decay τ1 and τ2 (ms) and amplitudes A1 and A226.
- Morphometric measurements
- Mount a clean, glass slide on the experimental bath chamber. Coat the slide with 2-3 µL of laminin and allow it to dry for 30 s before pouring ~400 µL of the fiber suspension onto the slide. Allow the fibers to adhere to the laminin for 10-15 min at room temperature.
- Evoke single twitches to verify the viability of the fibers by applying rectangular current pulses (0.8-1.2 ms) through the two platinum electrodes placed along either side of the experimental chamber. Even when attached to laminin, the contraction of the fibers is still visible mainly at the extremes.
- Acquire images of the alive fibers using 10x and 20x objectives and a camera of at least 5 megapixels mounted on an inverted fluorescence microscope. Store the images in .TIFF format for offline analyses.
NOTE: A set of ~2-6 images may be needed to completely capture a long fiber. - Image a microscope micrometer calibration ruler under the same magnification. Store the images in .TIFF format for offline analyses.
- Measure the lengths and diameters of the fibers using the calibration tool of free software for image analyses as follows:
- Establish a relation between pixels and the known distance (µm) in the images with the help of the microscope micrometer calibration ruler by using the Analyze/Set scale tool as shown in Supplemental Figure S1.
- Measure lengths once from one tip to the other of the fiber and diameters in 2-6 different places along the fiber (1-2 measurements per image, depending on its length), as in Supplemental Figure S1.
- Report the value of length (µm or mm) and the average of all diameters (µm) measured per fiber.
- Myosin heavy chain expression studies
NOTE: For details about the determination of MHC by immunofluorescence43 and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE)33,44,45,46 in whole muscles, please see Supplemental File 1. The protocol for fiber typing by immunofluorescence determination of MHC in the suspension of FDB-isolated fibers is as follows:- Coat each of five clean, glass slides with 2-3 µL of laminin and allow it to dry for 30 s before pouring ~300 µL of the fiber suspension onto each slide. Allow the fibers to adhere to the laminin for 4 h at room temperature.
- Fix the preparations with freezer-cooled acetone for 30 min at room temperature.
- Wash gently 3x with PBS.
- Permeabilize the cell membranes with PBS supplemented with 0.7% Triton X-100 for 15 min at room temperature.
- Wash gently 3x with PBS supplemented with 0.2% bovine serum albumin (BSA) and 0.04% Triton X-100, and subsequently block with PBS with 2% BSA, 2% goat serum, and 0.4% Triton X-100 for 30 min at room temperature.
- Wash gently 3x with PBS supplemented with 0.2% BSA and 0.04% Triton X-100 and incubate with the primary antibodies as follows:
- Dilute each anti-MHC primary antibody in a separate vial in PBS with 1% BSA and 0.04% Triton X-100: anti-I (1:1,500), anti-II (1:600), anti-IIA (use entire conditioned media from the hybridoma), and anti-IIB (1:500).
- Incubate each slide with one antibody and the remaining slide with PBS as a control for 12-16 h at 4 °C.
NOTE: In this protocol, fibers type IIX remained unlabeled in all samples.
- Wash gently 3x with PBS and incubate all slides with the secondary antibody (1:800) coupled to a fluorescent green molecule for 1-2 h at room temperature.
- Stain nuclei with 1 µg/mL Hoechst for 15 min.
- Wash gently 3x with PBS, carefully add 20-40 µL of mounting medium, and place a coverslip.
NOTE: Gentle solution exchanges and washing ensure that dozens of fibers remain attached to the slide, making the experiment statistically sound. - Visualize each slide using a 10x objective suitable for fluorescence and a filter set with the following wavelengths for excitation/dichroic/emission: 450-490/510/515 nm and count all positive and negative fibers. Alternatively, acquire fluorescence images using the same technical conditions and a camera of at least 5 megapixels mounted on an inverted fluorescence microscope and store them in .TIFF format for offline analyses.
- Record the positive and negative fibers of each slide in a database and calculate the percentages of positive I, IIA, IIB, and total II fibers based on the total number of fibers present in the corresponding slide. Calculate the percentage of IIX fibers by subtracting the sum of IIA+IIB from the percentage of total II fibers. Estimate the percentage of hybrid I/IIA fibers by subtracting the sum of I+II from a value of 100%. Finally, subtract the percentage of hybrid cells from the total of I and II to have the pure type I and II fibers.
NOTE: In MHC composition studies, fiber types are designated by a capital letter while isoforms are designated by a lowercase letter46.
- Hematoxylin and eosin staining
- Coat a clean, glass slide with 2-3 µL of laminin and allow it to dry for 30 s before pouring ~300 µL of the fiber suspension onto the slide. Allow the fibers to adhere to the laminin for 4 h at room temperature.
- Fix the preparation with Carnoy´s solution (60% absolute ethanol, 30% chloroform, 10% acetic acid) for 5 min at room temperature.
- Incubate with hematoxylin for 90 s.
- Wash gently 3x with tap water.
- Incubate with 1% eosin Y prepared in 70% ethanol for 30 s.
- Wash gently 3x with tap water.
- Immerse 3x into absolute ethanol.
- Incubate in xylol for 60 s.
- Add 20-40 µL of mounting medium and visualize with a conventional microscope. Acquire images at the desired magnification using a color camera of at least 5 megapixels.
5. Statistical analyses and graphing
NOTE: The experimental unit is a muscle fiber.
- Express results as mean ± standard deviation and calculate confidence intervals 95% (CI95%) for some analyses.
- To compare length, diameter, and Ca2+ transients´ kinetics among groups, perform analysis of variance (ANOVA) and post-hoc tests with the correction of Bonferroni.
- Assess normality and variance equality using the Shapiro-Wilk and Levene's tests, respectively.
- Consider the differences significant when p < 0.05.
Sarcoplasmic Ca2+ concentration during a twitch
To demonstrate the feasibility of physiological experiments in the set of dissociated fibers and to extend our previous findings on excitation-contraction coupling (ECC) and fiber types, Ca2+ transients were acquired in fibers from all muscles. First, FDB (n = 5) and EDL (n = 7) showed Ca2+ kinetics known as morphology type II (MT-II). These are fast, spiky signals, whose RT lasts ~1 ms; its decay phase can be fitted with a biexponential function with the first component (A1) larger than 30% of the whole amplitude, and its peak [Ca2+] is between 15 and 30 µM33,47 (Figure 3B). Their results are pooled (n = 12) in the first column of Table 1, while the results of the soleus´ Ca2+ transients (n = 6) are presented in the second column of Table 1. The soleus´ signals were classified as morphology type I (MT-I, Figure 3B) -wider, with an RT over 1.2 ms, an A1 lower than 30%, and a peak [Ca2+] between 7 and 13 µM33,47. These two columns were regarded as references for comparing the Ca2+ transients of the new muscles. Since all fibers from PDQA (n = 4), PL (n = 6), and EHL (n = 4) shared the MT-II (Figure 3B), their data are pooled (n = 14) in the third column of Table 1. These signals showed an average RT of ~1 ms, an A1 of ~45%, a peak [Ca2+] over 15 µM, and compare very well with the results presented in the first column but clearly differ from those shown in the second column, as confirmed by the statistical analysis (Table 1). The fastest signal of the whole sample came from an EHL fiber, with a ΔF/F of 0.66, [Ca2+] of 16.99 µM, RT of 0.85 ms, HW of 2.42 ms, and DT of 10.56 ms. τ1 and τ2 were 1.63 and 7.21 ms, respectively, while A1 and A2 values were 56.60% and 43.40%. The slowest signal came from a soleus fiber, with a ΔF/F of 0.41, [Ca2+] of 9.76 µM, RT of 1.56 ms, HW of 9.43 ms, and DT of 31.88 ms. τ1 and τ2 were 2.81 and 96.42 ms, respectively, while A1 and A2 values were 19.55% and 80.45%.
A battery of short, intermediate, and long fibers
The observation of clear differences among the fibers according to their muscle source enabled a more complete morphometric characterization. The histograms of Figure 4 show striking variations in the lengths of the fibers across all muscles. This is highlighted when comparing the shortest fiber from FDB (227.06 µm) with the longest one from soleus (5.69 mm). The mean length values are summarized in Table 2. There were significant statistical differences among groups (p < 0.01). These results allow for the classification of FDB fibers as short (<1 mm); PDQA and PL as intermediate (1 to 3 mm); and EDL, EHL, and soleus as long (>3 mm) (Supplemental Figure S2).
Conversely, minor differences were observed in the average diameters of the fibers of all muscles (Table 2) and the distribution of the values (Figure 4). Still, there were significant statistical differences among groups (p < 0.01). When evaluated in detail, there were differences between FDB and each other muscle and between FDB (FDB 38.40 ± 9.40 µm, n = 370) and the pool of intermediate and long fibers (45.07 ± 9.99 µm, n = 422, p < 0.05). The thinnest cell of the whole sample measured 18.42 µm (FDB) and the thickest reached 82.79 µm (PDQA).
Fiber types used in physiological experiments
First, fiber types present in each whole muscle were determined by immunofluorescence. Except for the soleus, the muscles showed a predominance of over 76% of type II fibers (Supplemental Figure S3, Table 3, and Table 4). EHL, PL, and PDQA are particularly fast muscles, with over 90% of fast fibers and up to 58.8% of type IIB fibers, as found in PDQA. EHL and PDQA were virtually devoid of type I fibers. Hybrid I/IIA fibers were present in all muscles in low percentages. As expected, type I and IIA fibers accounted for over 82% of the total fibers of the soleus. This muscle is almost devoid of the fastest IIB fibers. According to the profile of fiber types, the soleus is the slowest and the PDQA is the fastest muscle of the six analyzed.
Thereafter, and because it is more relevant for single-fiber physiological experiments, the question of which types of fibers appear in the cell suspension derived from the dissociated FDB was addressed. It was found that the profile of fibers in the cell suspension reflects the composition of fibers present in the cryosections: three out of four dissociated fibers were type II (Figure 5 and Table 3).
Finally, the composition of the muscles was confirmed by separating their MHC isoforms through SDS-PAGE. The results were consistent with the general pattern observed in the immunofluorescence assays. The soleus was enriched in MHC IIa and I, while the EDL, EHL, and peronei were enriched in MHC IIx and IIb but were devoid of I (Supplemental Figure S4).
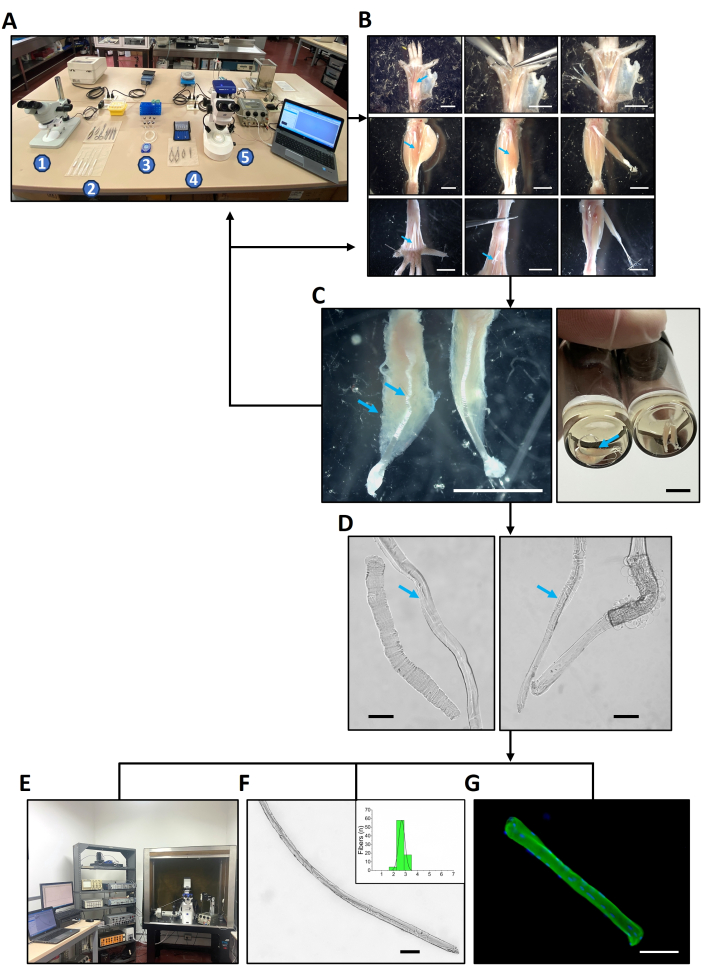
Figure 1: Design of the study. (A) Equipment and solutions to be set up before starting the dissection procedure. 1. Stereoscope number one. 2. From bottom to top: a set of five fire-polished pipettes, dissection tools, and collagenase vials inside a portable cooler (yellow case). 3. Filter case, dissection chamber, labeled glass vials with cap, rack with filtered solutions. 4. Dissection tools. 5. Stereoscope with dissection chamber numbers two coupled to a camera and an electric stimulator. (B) The dissection procedure of the set of six hindlimb muscles of mice as further explained in Figure 2. (C) After dissection, muscle contraction and integrity must be verified before continuing to the next protocol step. The left muscle in the left panel shows a wavy, hypercontracted, unresponsive PL muscle (blue arrow), which shall not be used to obtain dissociated fibers. Instead, the dissection protocol needs to be optimized and started again. The well-dissected PL counterpart (right muscle in the left panel) displays an elongated appearance and visibly contracts (Supplemental Video S1). The panel on the right shows two EDL (blue arrow) and two FDB muscles within the collagenase solution with the correct, straight appearance. (D) Once the muscles are dissociated, pour the isolated fibers into the experimental chamber and check their contraction and integrity. On the left panel, live (blue arrow) and dead PL fibers. On the right panel, live (blue arrow) and dead EDL fibers. (E–G) Three sets of experiments were performed with the isolated fibers: (E) Sarcoplasmic Ca2+ concentration measurements during a twitch (results in Figure 3). (F) Morphometric analyses (results presented in Figure 4). This example shows a PL fiber. (G) Immunofluorescence for myosin heavy chain expression studies (results illustrated in Figure 5). This example shows an isolated FDB fiber. Scale bars = 5 mm (B,C), 100 µm (D,F,G). Abbreviations: FDB = flexor digitorum brevis; PL = peroneus longus; EDL = extensor digitorum longus. Please click here to view a larger version of this figure.
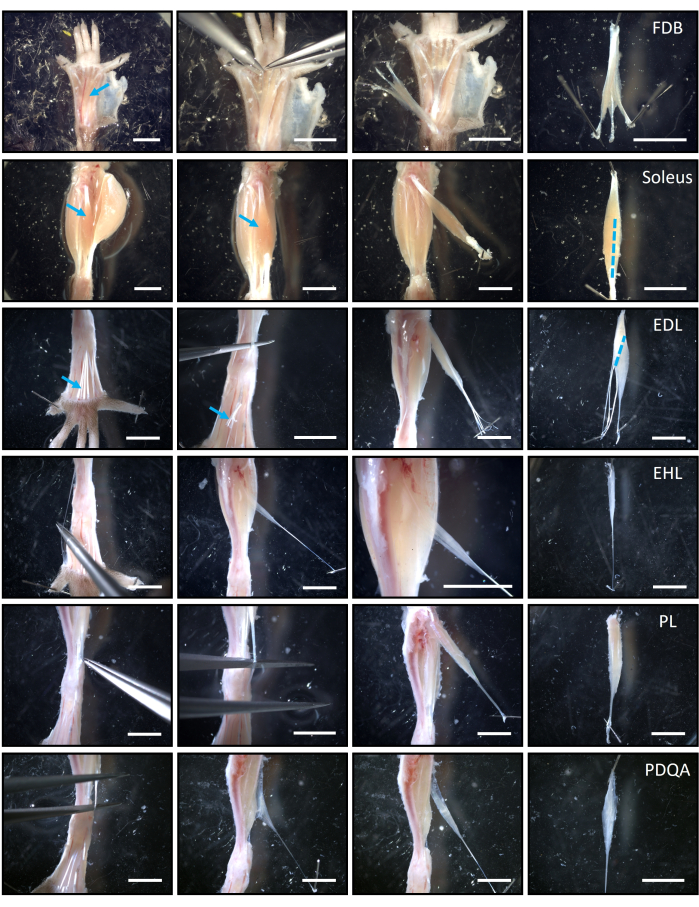
Figure 2: Anatomical references and general procedure to dissect the set of six hindlimb muscles of mouse. Rows, from top to bottom, present the muscles according to the best order of dissection: FDB, soleus, EDL, EHL, PL, and PDQA. Columns, from left to right, present the milestones during dissection. The first column shows the muscles or their distal tendons exposed so the dissection can start. For instance, blue arrows point to the FDB and soleus in situ and to the distal tendons of the EDL. The following two columns illustrate the dissection itself and the muscles exposed after the distal tendon(s) is/are cut, as labeled, for example, with the blue arrow in the EDL row. It is recommended that the branch of the FDB directed to the fifth digit is removed. The rightmost column presents the muscles once completely removed, with the proximal tendons oriented to the upper part of the images. Blue, discontinuous lines (extreme right column) illustrate the longitudinal or diagonal cuts that need to be performed in the soleus and EDL muscles to ensure that the collagenase enters their bulk. Scale bars = 5 mm. Abbreviations: FDB = flexor digitorum brevis; PL = peroneus longus; EDL = extensor digitorum longus; EHL = extensor hallucis longus; PDQA = peroneus digiti quarti. Please click here to view a larger version of this figure.
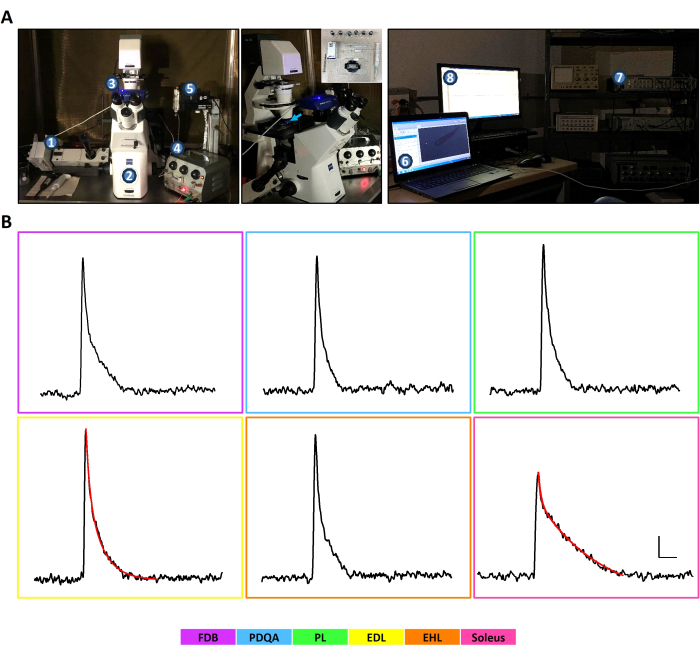
Figure 3: Ca2+ transients recorded in fibers obtained from a set of six hindlimb muscles of mouse. (A) Setup used for the acquisition of Ca2+ transients under dimmed illumination. Left panel: 1. PMT connected to an inverted fluorescence microscope (2). 3. Camera coupled to the microscope. 4. Electric stimulator coupled to the experimental chamber. 5. The micromanipulation system can be used when electrophysiology and injection assays are planned to complement the Ca2+ transients acquisition. Middle panel: the experimental chamber (insert) with the loaded cells is mounted onto the stage of the microscope and illuminated with blue light (450-490 nm, blue arrow) to excite the dye. In this setup, items 1 to 5 are placed over an antivibration table and inside a Faraday´s cage. Right panel: Once the quality of contraction and dye loading is verified with the camera (6), the light emitted is directed to the PMT, the electrical stimulation starts, and the signal is fed to the digitizer (7) and to the acquisition software (8) to record the Ca2+ transient. The integrated function of these elements during a live experiment can be seen in Supplemental Video S6. (B) Representative, calibrated Ca2+ transients of FDB, PDQA, PL, EDL, EHL, and soleus fibers of mouse. The similar, rapid, narrow signals of FDB, PDQA, PL, EDL, and EHL confirm that they have MT-II already known to derive from IIX and IIB fibers, which are the most abundant in these muscles. In contrast, the soleus transient is slower and smaller, typical of MT-I, originally described in I and IIA fibers. The red curve over the EDL and soleus signals demonstrates that the decay phase of MT-I and MT-II can be fitted with a biexponential decay function. Calibration bars apply to all panels. Vertical bar = 2.5 µM of [Ca2+], horizontal bar = 10 ms. The color key at the bottom helps identify the muscles. Abbreviations: FDB = flexor digitorum brevis; PDQA = peroneus digiti quarti; PL = peroneus longus; EDL = extensor digitorum longus; EHL = extensor hallucis longus; PMT = photomultiplier tube; MT-I = morphology type I; MT-II = morphology type II. Please click here to view a larger version of this figure.
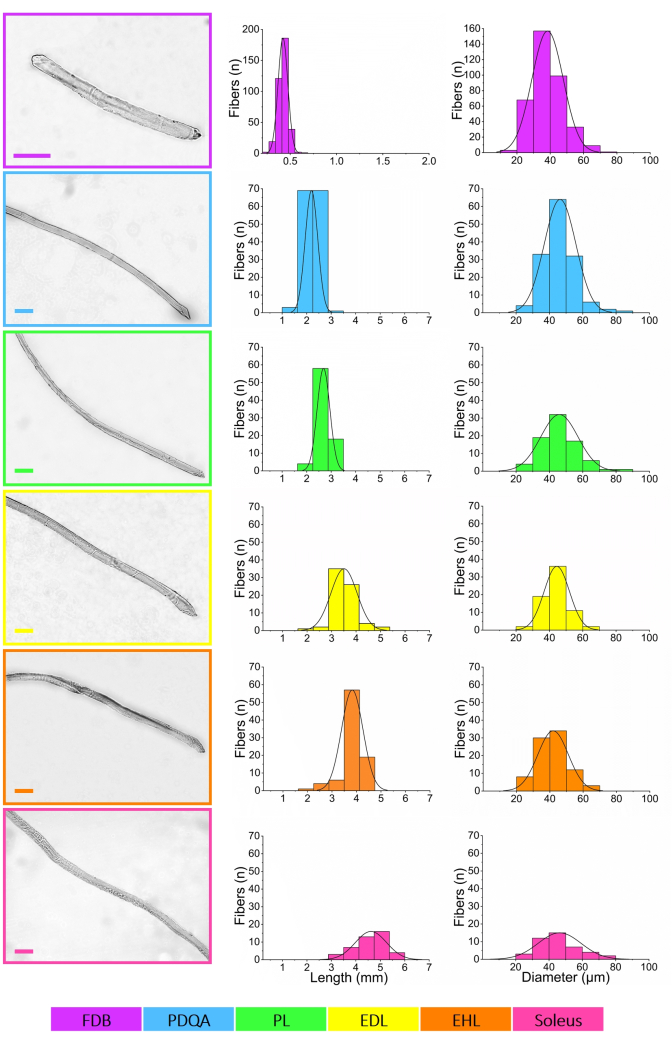
Figure 4: Morphometric measurements of the short, intermediate, and long fibers obtained by enzymatic dissociation of the set of six hindlimb muscles of mouse. Intact, healthy, representative fibers from each muscle are depicted in the leftmost column panels. Images were edited only to reduce background noise and enhance contrast, with no direct manipulation of the muscle fibers. Scale bars = 100 µm (all panels). The measurement protocol, as well as the appearance and quality of the complete long fibers, can be further viewed in Supplemental Figure S1. Middle column shows the distributions of lengths, ordered from the muscle that rendered the shortest fibers (FDB), to the one with the longest fibers (soleus). There is a continuum of lengths so that there is some overlap between muscles, spanning a total of ~5.5 mm. Diameter distributions are presented in the rightmost column panels. Most fibers are between 30 and 60 µm, with small differences among muscles. Test-retest analyses (n = 78 fibers, equivalent to 9.85% of the whole sample of n = 792) of the morphometric measurements showed very high reproducibility (mean coefficient of variation of 1.77%─confidence interval 95%, CI95%, 1.26-2.27%─mean coefficient of correlation of 0.99 (CI95% 0.98-0.99)), highlighting the good reliability of the results. Gaussian distribution curves were added with graphing licensed software. Supplemental Figure S2 shows bar plots of the mean values of length and diameter and the merges of length and diameter distributions of all muscles for easier comparison. Color key at the bottom helps identify the muscles. Abbreviations: FDB = flexor digitorum brevis; PDQA = peroneus digiti quarti; PL = peroneus longus; EDL = extensor digitorum longus; EHL, = extensor hallucis longus. Please click here to view a larger version of this figure.
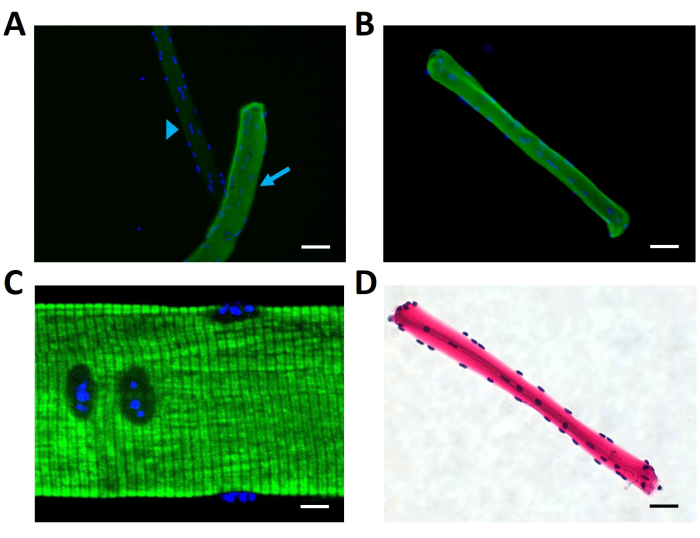
Figure 5: Immunofluorescence assays for fiber types in the cell suspension in dissociated FDB muscles. Images of different fields show intact, dissociated FDB fibers adhered to the bottom of the slides. (A) An antimyosin II-positive fiber (green fluorescence, diagonal light blue arrow) clearly contrasts with a negative one (light blue arrowhead), demonstrating the discrimination power of the assay. The presence of both fibers in the image can be confirmed because of the labeling of the nuclei (blue fluorescence). (B) A different field shows another antimyosin II-positive fiber. (C) The correct identification of the myosin heavy chain in the A bands (green fluorescence) by the antibodies was confirmed using confocal microscopy. The most notorious transversal black striations correspond to the I bands, enriched in actin, and thus are expected not to be recognized by the antibodies. (D) Hematoxylin labels in purple the typical peripheral, abundant nuclei, and eosin labels the sarcoplasm of the fiber, showing an intact, mature cell. Scale bars = 50 µm (A,B,D); 10 µm (C). Abbreviation: FDB = flexor digitorum brevis. Please click here to view a larger version of this figure.
| Fibers | FDB and EDL | Soleus | PDQA, PL and EHL | p |
| n | 12 | 6 | 14 | |
| ΔF/F | 0.68±0.15 | 0.46±0.06*,† | 0.66±0.12 | <0.01 |
| [Ca2+] (µM) | 17.46±5.18 | 11.14±1.86*,† | 16.82±3.29 | <0.01 |
| RT (ms) | 0.95±0.10 | 1.26±0.20*,† | 0.95±0.09 | <0.01 |
| Rise slope (F/ms) | 5.97±1.41 | 3.12±0.79*,† | 5.83±0.97 | <0.01 |
| HW (ms) | 3.59±0.85 | 8.17±3.00*,† | 3.19±0.54 | <0.01 |
| DT (ms) | 16.98±7.37 | 27.26±7.13*,† | 11.32±2.06* | <0.01 |
| Decay slope (F/ms) | -0.24±0.07 | -0.10±0.03*,† | -0.35±0.08* | <0.01 |
| τ1 (ms) | 1.86±0.37 | 2.07±0.66† | 1.57±0.18 | <0.05 |
| τ2 (ms) | 11.50±3.93 | 46.38±27.14*,† | 8.29±2.14 | <0.01 |
| A1 (%) | 48.14±7.27 | 25.94±8.98*,† | 44.59±8.29 | <0.01 |
| A2 (%) | 51.86±7.27 | 74.06±8.98*,† | 55.41±8.29 | <0.01 |
| Morphology | MT-II | MT-I | MT-II |
Table 1: Ca2+ transients' kinetics in enzymatically dissociated fibers obtained from a set of six hindlimb muscles of mouse. Values are mean ± standard deviation. p values correspond to the Analysis of variance test. *Significantly different from the FDB and EDL groups. †Significantly different from the PDQA, PL, and EHL groups. Abbreviations: F = fluorescence; [Ca2+] = cytosolic peak Ca2+ concentration; RT = rise time; HW = duration at half maximum; DT = decay time; τ1 and τ2 = time constants of decay; A1 and A2 = amplitudes of decay; FDB = flexor digitorum brevis; PDQA = peroneus digiti quarti; PL = peroneus longus; EDL = extensor digitorum longus; EHL = extensor hallucis longus; MT-I = morphology type I; MT-II = morphology type II.
| Fibers | FDB | PDQA | PL | EDL | EHL | Soleus | p |
| n | 370 | 142 | 80 | 70 | 87 | 43 | |
| Length (mm) | 0.42±0.05* | 2.20±0.26** | 2.69±0.26† | 3.51±0.53†† | 3.83±0.44‡ | 4.62±0.64 | <0.01 |
| Diameter (µm) | 38.40±9.40* | 46.39±9.50¶ | 45.98±11.18 | 44.43±7.62 | 41.96±9.16 | 46.36±12.85 | <0.01 |
Table 2: Morphometric measurements of the short, intermediate, and long fibers obtained by enzymatic dissociation of the set of six hindlimb muscles of mouse. Values are mean ± standard deviation. p values correspond to the Analysis of variance test. *Statistically different from PDQA, PL, EDL, EHL, and soleus. **Significantly different from PL, EDL, EHL, and soleus. †Significantly different from EDL, EHL, and soleus. ††Significantly different from EHL and soleus. ‡Significantly different from soleus. ¶Significantly different from EHL. Abbreviations: FDB = flexor digitorum brevis; PDQA = peroneus digiti quarti; PL = peroneus longus; EDL = extensor digitorum longus; EHL = extensor hallucis longus.
| Muscle/Fibers | N | n | Total type I + I/IIA (%) | Total type II (%) |
| FDB | 5 | 747 | 21.94±11.47 | 78.06±11.47 |
| *FDB | 8 | 1483 | 23.46±2.51 | 76.54±2.51 |
Table 3: Fiber types in cryosections and in the cell suspension in dissociated FDB muscles of mouse. Values are mean ± standard deviation. N refers to the number of animals, while n refers to the total number of fibers analyzed in the experiments. FDB row presents data of the whole muscle as determined in cryosections, while *FDB row corresponds to isolated fibers in the cell suspension after dissociating the muscle. The "Total type II" column reflects pure fibers type IIA, IIX, and IIB. Abbreviation: FDB = flexor digitorum brevis.
| Muscle | N | n | Type I (%) | Type I/IIA (%) | Type IIA (%) | Type IIX (%) | Type IIB (%) | Total type II (%) |
| PDQA | 4 | 597 | 0.00±0.00 | 10.15±2.62 | 19.33±6.19 | 11.68±7.57 | 58.84±7.63 | 89.85±2.62 |
| PL | 4 | 576 | 3.44±3.94 | 2.04±2.48 | 23.01±7.03 | 35.85±5.66 | 35.66±9.36 | 94.52±3.90 |
| EDL | 4 | 826 | 0.00±0.00 | 10.44±8.33 | 5.67±5.07 | 24.75±10.93 | 59.15±11.36 | 89.56±8.33 |
| EHL | 5 | 618 | 0.24±0.76 | 5.29±3.31 | 19.04±6.83 | 21.18±10.91 | 54.25±11.73 | 94.47±3.05 |
| Soleus | 4 | 729 | 32.18±4.48 | 1.74±1.30 | 50.37±7.46 | 14.58±6.94 | 1.12±1.93 | 66.08±5.59 |
Table 4: Fiber types in cryosections of the muscles which give intermediate and long fibers of mouse. Values are mean ± standard deviation. N refers to the number of animals, while n refers to the total number of fibers analyzed in the experiments. All rows present data of the whole muscle as determined in cryosections. Type I, IIA, IIX, and IIB columns reflect pure fibers, while the I/IIA column refers to hybrid fibers. The "Total type II" column is the summation of the type IIA, IIX, and IIB columns. Abbreviations: PDQA = peroneus digiti quarti; PL = peroneus longus; EDL = extensor digitorum longus; EHL = extensor hallucis longus.
Supplemental File 1: Myosin heavy chain expression studies in whole muscles. The protocols present details for the determination of myosin heavy chain isoforms by immunofluorescence in cryosections and by sodium dodecyl sulfate-polyacrylamide gel electrophoresis in homogenates of the whole muscles. Please click here to download this File.
Supplemental Figure S1: Details of the morphology of the fibers obtained by enzymatic dissociation of the set of six hindlimb muscles of mouse. (A) The microscope micrometer calibration ruler open to set the scale in the image analyses software is shown on the left. The right panel shows how the diameter was repeatedly measured as indicated by the blue lines perpendicular to the long axis of the fiber (blue arrows), while the length was measured once from tip to tip as illustrated by the blue line along the major axis of the fiber. (B) Several images were assembled with minor editing to demonstrate the long and healthy appearance of PDQA, PL, EDL, EHL, and soleus fibers (small blue, green, yellow, orange, and pink inserts). The assembled images were further edited to give the fibers a better look (large, black, and white images). (C) DIC images of FDB, PDQA, PL, and soleus fibers highlighting the normal irregularity of their tip and their well-known transversal striations. DIC appearance of the remaining EDL and EHL fibers can be seen in Supplemental Video S3 and Supplemental Video S4. Scale bars = 100 µm. The color key at the bottom helps identify the muscles. Abbreviations: FDB, flexor digitorum brevis; PDQA, peroneus digiti quarti; PL, peroneus longus; EDL, extensor digitorum longus; EHL, extensor hallucis longus; DIC = Differential interference contrast. Please click here to download this File.
Supplemental Figure S2: Bar plots and merged length and diameter distributions of the fibers obtained by enzymatic dissociation of the set of six hindlimb muscles of mouse. (A) Bar plots (mean ± standard deviations) and (B) merged histograms confirm the disparity of the lengths but the similarity of the diameters across all groups. *Significantly different from PDQA, PL, EDL, EHL, and soleus. **Significantly different from PL, EDL, EHL, and soleus. †Significantly different from EDL, EHL, and soleus. ††Significantly different from EHL and soleus. ‡Significantly different from soleus. ¶Significantly different from EHL. The color key at the bottom helps identify the muscles. Abbreviations: FDB = flexor digitorum brevis; PDQA = peroneus digiti quarti; PL = peroneus longus; EDL = extensor digitorum longus; EHL = extensor hallucis longus. Please click here to download this File.
Supplemental Figure S3: Immunofluorescence studies for fiber type composition of the set of six hindlimb muscles of mouse. Representative antibody-labeled cryosections used for the analyses demonstrate the good quality and intactness of the samples. There is a clear predominance of fiber type II in all muscles, except for the soleus in which the amount of fiber type I is sizeable. Scale bars = 100 µm. The color key at the bottom helps identify the muscles. Abbreviations: FDB = flexor digitorum brevis; PDQA = peroneus digiti quarti; PL = peroneus longus; EDL = extensor digitorum longus; EHL = extensor hallucis longus. Please click here to download this File.
Supplemental Figure S4: Electrophoretic separation of MHC isoforms in the set of six hindlimb muscles of mouse. (A) Two representative complete gels run on different days with different samples demonstrate the quality, cleanness, and reproducibility of the separation and migration distance of the MHC isoforms when stained with Coomassie blue. Although these two images were slightly edited to enhance contrast and clarity for the reader, the analyses were performed in unedited gels. (B) Examples of the analyses of the bands for each of the six muscles. After an ROI is selected in the lanes of the gel, a plot profile is generated and a Gaussian fit is used to estimate the proportion of MHC isoforms in the set of six muscles. The MHC composition found was: FDB: I 9.0 ± 5.0%, IIa + IIx 91.0 ± 5.0% (n = 3); PDQA: IIa + IIx 25.9 ± 2.4%, IIb 74.1 ± 2.4% (n = 3); PL: IIa + IIx 24.9 ± 2.2%, IIb 75.1 ± 2.2% (n = 3); EDL: IIa + IIx 22.0 ± 2.3%, IIb 78.0 ± 2.3% (n = 4); EHL: IIa + IIx 27.5 ± 1.0%, IIb 72.5 ± 1.0% (n = 3); soleus: I 35.8 ± 2.5%, IIa (+IIx + IIb when present) 64.2 ± 2.5% (n = 4). The color key at the bottom helps identify the muscles. The gray rectangle below the rightmost gel in A refers to the molecular weight marker indicating the migration of the IIa to ~ 200 KDa. Abbreviations: FDB = flexor digitorum brevis; PDQA = peroneus digiti quarti; PL = peroneus longus; EDL = extensor digitorum longus; EHL = extensor hallucis longus; MHC = myosin heavy chain; ROI = region of interest. Please click here to download this File.
Supplemental Video S1: The damaged, wavy, peroneus longus (PL, left) does not contract upon stimulation, while the intact PL (right) contracts well, even when it is farther from the electrodes. 0.8x magnification. The stimulation protocol releases rectangular current pulses of 1.2 ms, 0.7 Hz, and 50 V through two electrodes. Please click here to download this Video.
Supplemental Video S2: Contracting peroneus longus fiber. 20x magnification. Differential interference contrast mode. The stimulation protocol releases rectangular current pulses of 1.2 ms, 0.5 Hz, and 50 V through two platinum electrodes. Please click here to download this Video.
Supplemental Video S3: Contracting extensor digitorum longus fiber. 20x magnification. Differential interference contrast mode. The stimulation protocol releases rectangular current pulses of 1.2 ms, 0.5 Hz, and 50 V through two platinum electrodes. Please click here to download this Video.
Supplemental Video S4: Contracting extensor hallucis longus fiber. 20x magnification. Differential interference contrast mode. The stimulation protocol releases rectangular current pulses of 1.2 ms, 0.5 Hz, and 50 V through two platinum electrodes. Please click here to download this Video.
Supplemental Video S5. Contracting soleus fiber. 20x magnification. Differential interference contrast mode. The stimulation protocol releases rectangular current pulses of 1.2 ms, 0.5 Hz, and 50 V, through two platinum electrodes. Please click here to download this Video.
Supplemental Video S6: Live recording of a Ca2+ transient from an extensor digitorum longus fiber loaded with Mag-Fluo-4, AM as described in step 4.1.4. The stimulation protocol releases rectangular current pulses of 1.2 ms, 0.5 Hz, and 50 V through two platinum electrodes. Please click here to download this Video.
