Stage-specific MO injections provide a useful approach to analyze gene function in specific compartments of the embryo. Figure 1 presents a flow chart of the injection strategies used to test gene function in DFC/KV cells and how to introduce fluorescent beads to visualize fluid flow in KV. The distribution of fluorescent MO in successful stage-specific injected embryos is shown schematically in Figures 1D-F and in live embryos in Figure 2. An unsuccessful MO injection, in which the MO remains aggregated in the yolk cell, is shown in Figure 2D.
Successfully injected embryos-selected based on the localization of fluorescent MO-can be mounted at the 4-6 somite stages for delivery of fluorescent microbeads into the KV lumen (Figures 1G-H) to analyze fluid flow. Videomicroscopy is used to record bead movements in KV (Figures 1I-J), which can be analyzed qualitatively (Figures 3A-B) or quantitatively (Figures 3C-E) using ImageJ software. In a control embryo with normal flow, beads follow counterclockwise paths that can be visualized by making a maximum projection of fluorescent bead positions over time (Figure 3A) or by tracking individual beads over time (Figure 3C). To demonstrate loss of coordinated flow, we injected embryos with MO to knockdown Rho kinase 2b (Rock2b), which we have previously shown disrupts flow18. In Rock2b MO injected embryos, beads move randomly in KV (Figures 3B, D). ImageJ software was used to calculate the velocity of individual bead tracks. The average velocity of 5 beads from control and Rock2b MO embryos is shown in Figure 3E.
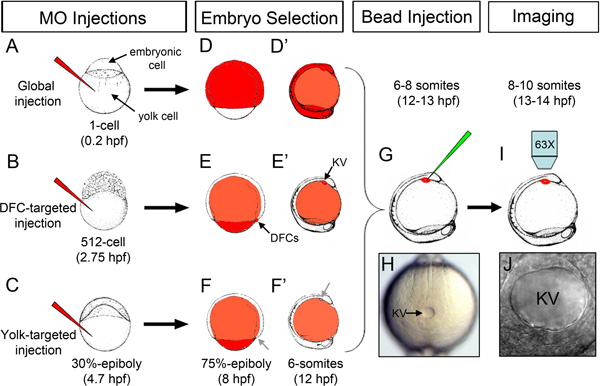
Figure 1. Overview of stage-specific MO injections and microbead injection into KV. (A) Injection of fluorescent MO (red) into the yolk at the 1-cell stage for global distribution of MO throughout the embryo. (B) DFC-targeted MO injection at the 512-cell stage to load MO into DFC/KV cells. (C) Yolk-targeted MO injection at 30%-epiboly stage to restrict MO to the yolk. (D-F’) Schematic representation of the distribution of fluorescent MO (red) in successfully injected embryos at the 75% epiboly stage (D, E, F) and the 6 somite stage (D’, E’, F’) following stage-specific injections. MO accumulates in the DFC/KV cell lineage when injected at the 512-cell stage (arrows in E, E’), but not when injected at 30% epiboly (arrows in F, F’). (G) Injection of fluorescent microbeads (green) into the KV lumen. (H) A properly mounted embryo with KV (arrow) facing up for microbead injection. (I) Imaging beads movement in KV using an upright microscope. (J) High magnification of an intact KV lumen injected with microbeads. Embryonic stages and embryo drawings are based on ref. 28.
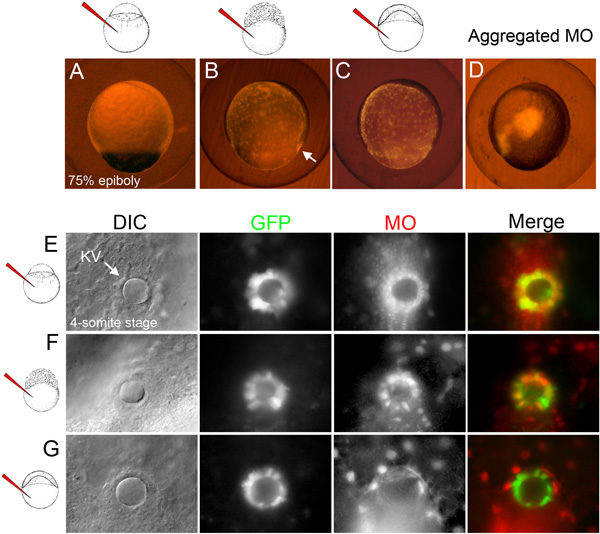
Figure 2. Selection of embryos following stage-specific MO injections. (A-C) Examples of selected embryos at the 75% epiboly stage (8 hpf) in which fluorescent MO (red) has either incorporated in all embryonic cells following global MO injection (A), diffused through the yolk and DFCs (arrow) following DFC-targeted MO injection (B) or remained in the yolk following yolk-targeted MO injection (C). (D) Example of an excluded embryo in which the fluorescent MO aggregated at the injection site. (E-G) Examples of fluorescent MO (red) distribution in selected embryos at the 4-somite stage (11.5 hpf). DIC images identify the KV lumen (arrow) in transgenic Tg(Dusp6:d2EGFP) embryos that express GFP in KV cells27. In the global MO injected embryo, MO was observed in KV and all surrounding cells (E). In the DFC-targeted MO injected embryo, MO co-localized with GFP in most KV cells and was also present in underlying yolk nuclei (F). In the yolk-targeted embryo, MO was found exclusively in yolk nuclei (G).
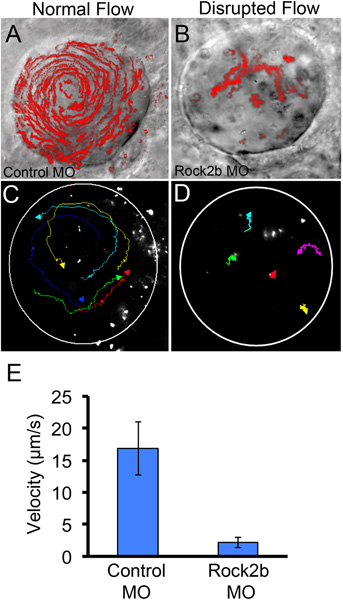
Figure 3. Qualitative and quantitative analyses of fluid flow in KV. (A-B) For qualitative analysis of flow, a maximum projection of a 10 sec movie showing movement of all fluorescent microbeads injected into KV has been superimposed on a DIC image of the KV lumen in a global control MO (A) or Rock2b MO (B) injected embryo. (C-E) To quantify flow, movement of individual microbeads (n=5) was tracked in a control embryo (C) and Rock2b MO injected embryo (D) and used to calculate an average bead velocity (E). Circles (C-D) approximate the KV lumen boundaries. Error bars (E) represent one standard deviation.