G protein-koblede reseptorer (GPCR) utgjør en av de største og mest mangfoldige familier blant alle celleoverflateproteiner. Deres tilstedeværelse i virveldyr, virvelløse dyr, planter, gjær og mugg, slim så vel som i protozoer og den tidligste diploblastic metazoa indikerer at GPCR er blant de eldste molekyler knyttet til signaltransduksjon 1.. Deres naturlige aktive ligander omfatter et bredt mangfold av eksterne stimuli inkludert peptider, biogene aminer, dufter, glykoproteiner, og fotoner to. Som sådan er disse reseptor-ligand-signalsystemer er involvert i mange forskjellige fysiologiske prosesser. Den brede funksjonelle spektrum gjør dem ideelt egnet for utvikling av terapeutiske medikamenter som dekker et bredt spekter av humane sykdommer. Om 50-60% av dagens narkotika mål er representert ved GPCRs 3,4. Foruten sin store betydning i den farmasøytiske industrien, GPCRs er også i søkelyset for utvikling av enny generasjon av artsspesifikke insekt 5,6 og plantevernmidler generelt. Fordi de naturlige ligander av mange GPCRs er fortsatt uidentifisert, klassifiseres de som foreldreløs GPCRs. Den deorphanization av disse reseptorene vil bedre forståelsen av deres fysiologiske roller i organismer og kan avdekke antatte mål for nye stoffet applikasjoner 7.
Siden den genomiske era, er det motsatte farmakologi strategi mye brukt for deorphanization av GPCR åtte. Denne metode innebærer at en orphan reseptorer er brukt som en "krok" til "fisk ut 'sin aktiverende ligand i et biologisk ekstrakt eller fra et bibliotek av syntetiske forbindelser. Den GPCR av interesse er derfor klonet og deretter transfektert i et mobiltelefon ekspresjonssystem. I de mest brukte metoder er reseptoraktivering bestemmes ved måling av endringer i konsentrasjonen av andre messenger molekyler 9 </sup>. De viktigste reseptor screening-analyser er avhengige av kalsium-sensitive bioluminiserende proteiner (f.eks, aequorin) 10 eller fluorescerende kalsium indikatorer (f.eks Fluo-4) 11. Den fluorescens-baserte analyser, hvori reseptor-uttrykkende celler blir lastet med en fluorescerende kalsiumindikator før ligand screening, har den fordel at de tillater high-throughput screening fordi de er enkle å bruke, kortlese tid, og fleksibiliteten til screening flere foreldreløse reseptorer på en enkelt plate 12.
Her blir fluorescens-baserte kalsium mobiliseringsanalyse grundig beskrevet og illustrert ved deorphanization prosessen av Drosophila melanogaster korte nevropeptid K (sNPF) reseptoren. Denne neuropeptidergic signalsystem ble opprinnelig karakterisert ved Mertens et al. i 2002 13 med en kalsium Bioluminescens analysen utført i kinesisk hamster eggstokk (CHO)14 og etter Feng et al. I 2003 med en elektrofysiologisk analyse ved hjelp Xenopus oocytter 15. Tilstedeværelsen av sNPF signaleringssystem ser ut til å være begrenset til rekken av Arthropoda, hvor det er innblandet i en rekke prosesser, inkludert regulering av foring, vekst, stressreaksjoner, bevegelse, og døgnrytme 16.
Forskning på neuropeptidergic signalanlegg i insekter kan ikke bare føre til nye mål for utvikling av insektmidler, men kjennskap til deres virkemåte kan også bli ekstrapolert til andre organismer så mange signalsystemer er generelt godt konservert gjennom evolusjon 17.. I det siste tiåret, har store fremskritt er gjort i deorphanization prosessen med insekt neuropeptidreseptorer GPCR. Til tross for disse tiltakene, har bare et lite antall reseptorer blitt matchet til deres beslektet ligand, og massevis av sekvensinformasjon fornye foreldreløs GPCRs har blitt tilgjengelig på grunn av den blomstrende av genomikk 18. Tilgjengeligheten av middels / høy throughput screening tilnærminger, som fluorescens-basert kalsium mobilisering analysen som har vist seg å være en mye brukt teknikk 9,18, er derfor uvurderlig.
Den fluorescens-baserte kalsium mobiliseringsanalyse som beskrevet her blir utført i den humane embryonale nyre 293T (HEK293T) cellelinje, og benytter et fluorescerende probe for å bestemme endringer i den intracellulære kalsiumkonsentrasjon ved reseptor-aktivering. For å sikre høy ekspresjon og oversettelses nivåer av reseptoren, en Kozak konsensussekvens 19 tilsatt til 5'-enden av den reseptor-kodende sekvens, som deretter klonet i en ekspresjonsvektor (for eksempel vektor pcDNA serie for pattedyr-cellelinjer). Ettersom det er vanskelig å forutsi den endogene G protein-kopling av en foreldreløse GPCR basert på sekvensinformasjonalene, de andre messenger molekyler (f.eks, kalsium eller camp) som er modulert etter reseptoraktivering ofte forblir ukjent før ligand identifikasjon. For å omgå dette problemet, promiskuøse G proteiner i G q familien (f.eks murine Gα 15 eller menneskelig Gα 16 [brukt her]) eller chimere G proteiner (f.eks, Gα qi5) som samhandler med de fleste GPCRs og indusere frigjøring av kalsium kan være co-uttrykt 20,21,22. Ved binding av ligand til dens reseptor, gjennomgår GPCR en konformasjonsendring som fører til aktivering av spesifikke intracellulære baner. Den guanosine diphosphate (BNP) molekyl, forpliktet i henhold til hvile på, til Gα 16 subenheten, vil bli erstattet av en guanosine trifosfat (GTP) molekyl. Dette frembringer en dissosiasjon av heterotrimeric G protein i en Gα 16 og Gβγ subenhet. Den Gα 16 subenheten aktiverer fosfolipase C &# 946; (PLCβ), som i sin tur hydrolyserer membran-bundet fosfatidylinositol bisfosfat (PIP 2) som resulterer i diacylglycerol (DAG) og inositol Trisphosphate (IP 3). IP 3 vil spre seg over hele cytoplasmaet og aktiverer IP 3-avhengige kalsiumkanaler tilstede i membranen av det endoplasmatiske retikulum, som induserer frigjøring av kalsium inn i cytoplasmaet.
Den kalsium-frigjøring ved reseptor aktivering skjer i løpet av sekunder, og kan bli detektert ved å laste celler før screening-analyse med et kalsiumsensitivt fargestoff, som Fluo-4-acetoksymetyl (AM) 11. Den AM ester-gruppen kan fluoroforen for å krysse cellemembranen og spaltes av ved cytoplasmatiske esteraser Inne i cellen. Følgelig er de negative ladninger av den fluorescerende fargestoff avslørt, og hindrer den fra å spre ut av cellen og slik at det å samhandle med kalsiumioner. Fluorescenssignalet of Fluo-4 er ubetydelig i celler under hvile på, bare inneholder kalsiumkonsentrasjonen i nanomolarområdet. Imidlertid, når kalsium er sluppet på reseptoraktivering, kan signalet øke konsentrasjonen avhengig til mer enn 100-ganger herved sikre et stort signal-til-støy-forhold. Fluo-4 oppviser dessuten et stort dynamisk område for rapportering [kalsium] rundt en K d (kalsium) på 345 nM, som gjør det egnet til å måle fysiologisk relevante kalsium endringer i et bredt spekter av celler. Den magnetisering av Fluo-4 skjer ved 488 nm og emisjon fluorescens måles ved 525 nm 11. Fluorimeters som fluorescens avbildningsplateleser (FLIPR) 23, Novostars eller FlexStation (stasjonsenhet) 12 er middels / høy gjennomstrømming systemer som tillater simultan forbindelsen tilsetningen og deteksjonen av den Fluo-4-signal ved reseptoraktivering for hver godt i en prøve-plate. Den kalsium mobiliseringsanalyse som beskrives her er avhengig av stasjonenEnheten 96-brønns mikroplate-systemet.
Den SoftMax Pro (software) blir brukt til å betjene stasjonen enheten samt for dataanalyse. Programmet viser umiddelbart resultater som grafer i 96-brønners format. Flere brønner kan velges samtidig for å sammenligne resultatet av disse brønner på den samme grafen. Verdier av brønner i hver kolonne Den relative fluorescerende enhet (RFU) blir samtidig målt i et tidsrom på to minutter, med start før tilsetning av forbindelsene til brønnene og fortsetter etter måling av det fluorescerende signal etter reseptoraktivering. Vanligvis, utviklingen av en agonist kurve er på linje med grunnlinjen til en aktiverende forbindelse ble tilsatt til cellene, noe som resulterer i en rask økning av det fluorescerende signal. Den topphøyde er korrelert med den endelige agonist-konsentrasjon i brønnen. Etter toppen faller fluorescenssignalet sakte mot baseline nivå. RFU målinger can bli omdannet til konsentrasjons-responskurver for å bestemme EC50-verdi (halv maksimal effektiv konsentrasjon) av en ligand. Generelt er i det minste tre uavhengige skjermer, hver med tre kopier av en konsentrasjonsserie, bør utføres for å sette sammen en pålitelig konsentrasjon-respons-kurve.
Det anbefales å inkludere flere positive og negative kontroller i eksperimentell design. Først av alt, en transfeksjon kontroll, det vil si innføringen av en reseptor med en kjent ligand, bør testes. Dette gjør det mulig å verifisere om transfeksjon agent var i drift. Innlemmelse av en kontrolleksperiment med en agonist for en endogen reseptor på cellelinjen, og en negativ kontroll (for eksempel vaskebuffer) er også anbefalt for å overvåke tilstanden og levedyktighet av cellene og for å utelukke muligheten for at vaskebufferen var forurenset med en faktor som kan lokke fram en auto-fluorescerende Whiteduft respons. Ofte brukte agonister er et peptid avledet fra protease-reseptor-1 (par 1), som fungerer som en PAR en selektiv agonist eller carbachol, som aktiverer den acetylcholin-reseptoren. Celler transfektert med tom ekspresjonsvektor bør også testes for å utelukke at aktive forbindelser kommuniserer med cellens endogene reseptorer. Optimalisering av flere parametere som er beskrevet i protokollen nedenfor, kan være nødvendig for ulike signalsystemer. En skjematisk figur av den komp fluorescens-baserte kalsium mobiliseringsanalyse er vist i figur 1.
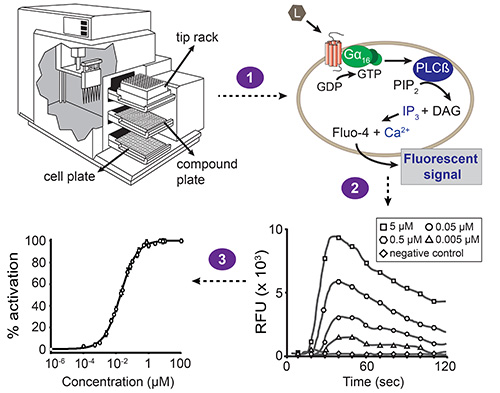
Figur 1. Generelle ordningen med fluorescens-basert kalsium mobilisering analysen. Automatisert væskehåndtering og samtidig fluorescens målinger er utført med stasjonenenhetsmikroplateleser, drevet av programvaren. Stasjonen enheten inneholder tre skuffer: en for cellen plate, sammensatte plate og tips rack. Den innebygde Pipetteenhet overføringer forbindelser fra én kolonne med det sammensatte plate til den tilsvarende kolonnen av cellene plate (trinn 1). Hver brønn av celleplate har et monosjikt av HEK293T celler som har blitt co-transfektert med GPCR av interesse og promiskuøse Gα 16 subenheten. Når en forbindelse som aktiverer reseptoren, er Gα 16-bundet GDP erstattet med GTP. Den Gα 16 subenheten deretter spaltes fra det Gβγ kompleks og aktiverer fosfolipase Cβ (PLCβ), som i sin tur hydrolyserer fosfatidylinositol bisfosfat (PIP 2) som resulterer i diacylglycerol (DAG) og inositol Trisphosphate (IP 3). IP 3 aktiverer IP 3-avhengige kalsiumkanaler tilstede i membranen av det endoplasmatiske retikulum, indusere frigjøring av kalsium into cytoplasma. Interaksjonen av kalsium med Fluo-4 (med hvilke cellene er lastet før sammensatte tillegg) resulterer i et fluorescerende signal (trinn 2). Programvaren presenterer resultatene som relative fluorescerende enhet (RFU)-verdier i funksjon av tiden, og topphøyder korrelerer med ligandkonsentrasjonen på en konsentrasjonsavhengig måte. Disse dataene kan deretter bli omdannet til en konsentrasjon-respons-kurve for å bestemme EC50 verdien av en ligand-reseptor-par (trinn 3).
