Drosophila Startle-induced Locomotion Assay
Wildtype, canton-S, flies showed a robust negative geotactic response with only approximately 88% and 5% of flies in the top and bottom sections respectively, of the double-vial apparatus after 30 sec (Figure 1). Flies exposed to 125 μM and 250 μM rotenone for 3 days showed a slight decrease in the number of flies in the top section and slight increase in the number of flies in the bottom section. Flies exposed to 500 μM rotenone showed significant defect in the negative geotactic response (p < 0.05 ANOVA, Bonferroni pair wise comparison) as evidenced by fewer flies in the top section and more flies in bottom section as compared to control flies (Figure 1). This defect in negative geotactic response due to inability to swiftly climb in the apparatus is indicative of a defect in startle-induced locomotion.
Drosophila Spontaneous Locomotion Assay
Wildtype, canton-S, flies showed 0.57 counts per minute as a measure of spontaneous mobility on the fourth day in the DAM (Figure 2). Flies exposed to 125 μM rotenone showed a similar level of spontaneous locomotion. By contrast, flies exposed to 250 μM and 500 μM rotenone showed approximately 50% lower measures (p < 0.05 ANOVA, Bonferroni pair wise comparison) of spontaneous locomotion (Figure 2). These flies moved at about 0.20 counts per minute, which is indicative of a rotenone-induced defect in spontaneous locomotion.
To account for initial discrepancies (if any) in locomotion not caused by rotenone exposure, we subtracted locomotion data between day 4 and day 3. Control flies showed an increase of about 0.1 counts per minute in spontaneous locomotion between days, while flies exposed to 125 μM rotenone exhibited a slight decrease of about 0.15 counts per minute (Figure 3). Flies exposed to 250 μM and 500 μM rotenone displayed more severe decreases in locomotion between days, with differences being approximately 0.3 and 0.5 (p < 0.05 ANOVA, Bonferroni pair wise comparison) counts per minute respectively. This data suggests a deficiency in spontaneous locomotion over time with exposure to rotenone and confirms single day analysis mentioned above–flies exposed to higher dosages of rotenone showed a decrease in spontaneous locomotion. Taken together, these methods reliably measure rotenone-induced deficiencies in spontaneous and startle-induced locomotion.
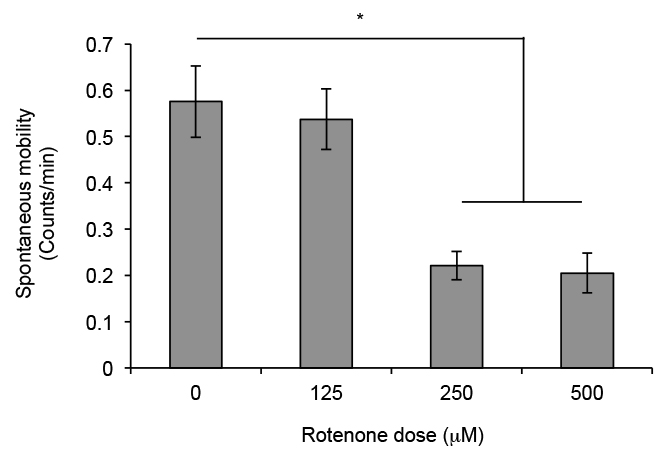
Figure 1. Startle-induced locomotion plot of flies exposed to increasing doses of rotenone. Wild type, canton-S, male flies were exposed to different dosages of rotenone for 3 days and surviving flies (8-12) were then tapped into the bottom of the double vial apparatus. Flies exposed to 500 μM rotenone show a significant decrease in the percent of flies in the top (A) and increase in the percent of flies in the bottom section (B) of the apparatus after 30 sec. This is indicative of a deficiency in startle response in flies exposed to rotenone. Columns represent the average percentage of 6 independent experiments. Error bars represent the standard error of mean; * p < 0.05 ANOVA, Bonferroni pair wise comparison.
Figure 2. Spontaneous locomotion plot of flies exposed to increasing doses of rotenone. Wild type, canton-S, male flies were exposed to different dosages of rotenone and counts per min on the fourth day after exposure were plotted. Counts were measured in a DAM. Flies exposed to 250 μM and 500 μM rotenone show a reduction in counts per min. This is indicative of a deficiency in spontaneous locomotion in flies exposed to rotenone. Columns represent the average counts per min on fourth day of 5 independent trials. Error bars represent the standard error of mean; * p < 0.05 ANOVA, Bonferroni pair wise comparison.
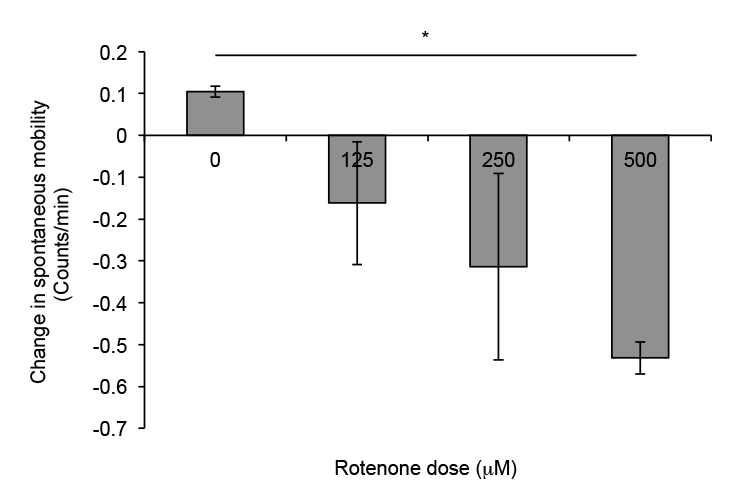
Figure 3. Change in spontaneous locomotion plot of flies exposed to increasing dosages of rotenone. Wild type, canton-S, male flies were exposed to different dosages of rotenone and the difference in counts per min on the third and fourth day after exposure were plotted. Counts were measured in a DAM. There is a dose dependent trend for decline in spontaneous locomotion with flies exposed to higher doses having a more negative change in locomotion. This is indicative of a decrease in movement. Columns represent the average change in locomotion per minute of 5 independent trials. Error bars represent the standard error of mean; * p < 0.05 ANOVA, Bonferroni pair wise comparison.
