After 1 hr of perfusion of the retinal preparations with oxygen-saturated standard solution (Figure 1A and B) ERG-amplitudes showed stabilization and less variation of amplitudes between single measurements. pH, osmotic pressure, temperature, and pO2 (except for hypoxia testing) were kept constant for all tests.
To isolate the photoreceptor signal from the signal of the inner retina, 1 mM aspartate was added to the standard solution to suppress the b-wave (Figure 1A). During testing of the effect of hypoxia, a decrease in a-wave amplitude of 87.0% was noted (p <0.01) after an exposition time of 45 minutes. At the end of the washout, a decrease of 36.5% was noted that was statistically significant (p = 0.03, Figure 2A). Additionally, an initial decrease in b-wave amplitudes of 87.23% was recorded, that equally reached statistical significance (p <0.01). In this setting a reduction of 25.5% was noted, that was statistically significant (p = 0.03, Figure 2B).
After exposition with 250 µm glutamate, a 7.8% non-significant reduction of a-wave amplitudes was detected after (p >0.05) the defined time interval. This was followed by a non-significant reduction of 1.9% (p >0.05, Figure 3A). Single measurements are shown in Table 1 and 2.
Concerning the b-wave, decreased amplitudes of the ERG by 83.7% were recorded that were statistically significant (p <0.01, Figure 3B). At the end of the washout, a b-wave recovery was noted resulting in a non-significant reduction of 2.3% after 75 minutes of perfusion with standard solution (p = 0.62).
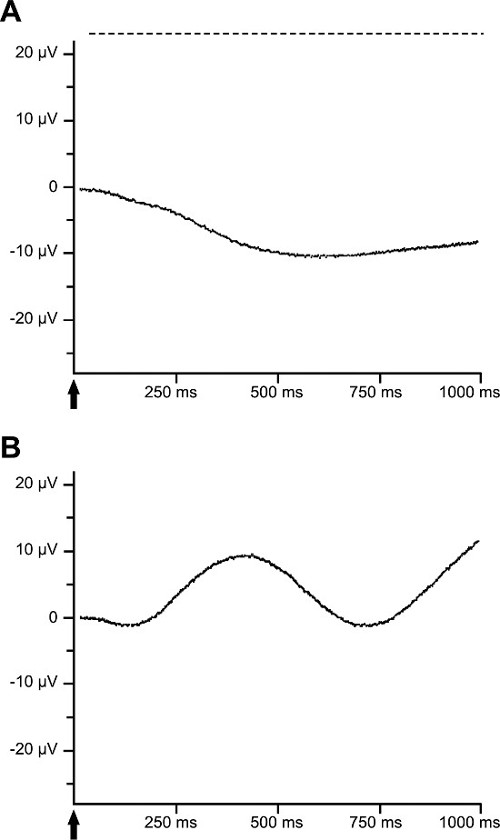
Figure 1: Example of an ERG measurement from the isolated perfused bovine retina. (A) The a-wave is shown in the ERG of the isolated perfused bovine retina. The b-wave is suppressed by adding 1 mM aspartate to the nutrient solution. (B) The b-wave is dominant under scotopic light conditions. A 10 ms light stimulus at a light intensity of 6.3 mlx is used.
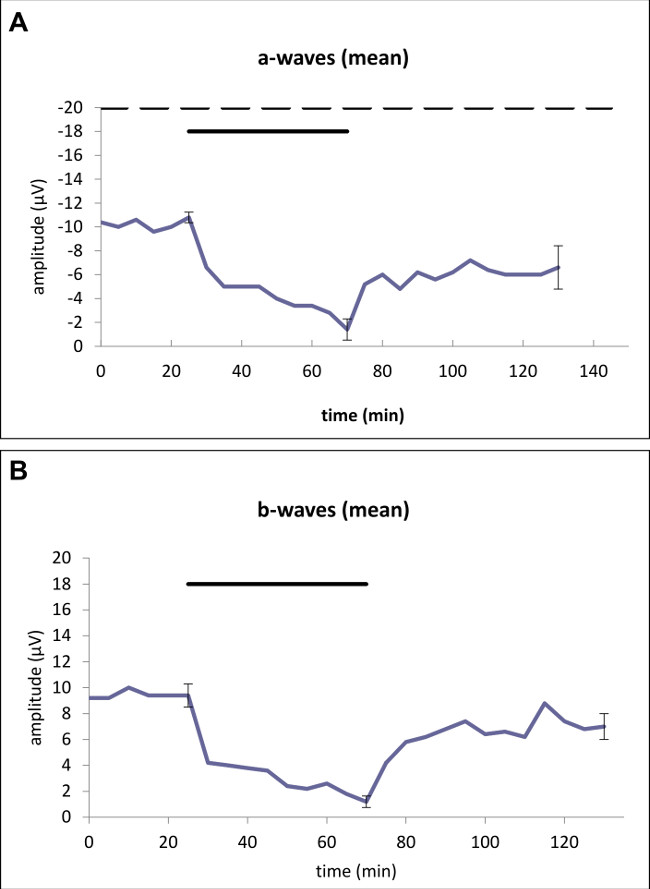
Figure 2: Effects of hypoxia after an exposure time of 45 min on the (A) a-wave and on the (B) b-wave amplitude of the ERG. Average of representative drug series (n = 5). The horizontal bar above the curve marks the hypoxia time. The dotted line (A) marks the application of aspartate 1 mM to unmask photoreceptor potential. Standard deviations for each series of experiments are given directly before and after application as well as at the end of the trial. Statistical analysis was performed at the time-points using the standard deviation (directly before and after application as well as at the end of the trial): (A) A decrease in a-wave amplitude of 87.0% was noted after an exposition time of 45 minutes compared to the beginning of the trial. At the end of the trial, a significant decrease of 36.5% was noted. (B) A significant decrease in b-wave amplitudes of 87.23% was recorded at the end of the exposition time. At the end of the trial, a significant reduction of 25.5% was noted.
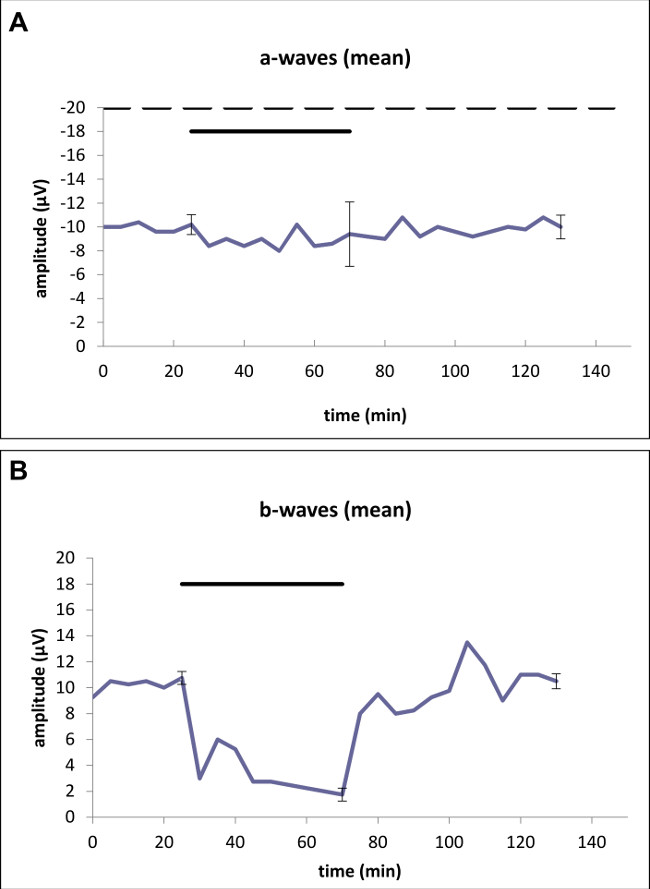
Figure 3: Effects of 250 µM glutamate applied for 45 min on the (A) a-wave amplitude and the (B) b-wave amplitude of the ERG of the isolated perfused bovine retina. Average of representative drug series (n = 5). The horizontal bar above the curve marks the application of glutamate. The dotted line (A) marks the application of aspartate 1 mM to unmask photoreceptor potential. Standard deviations for each series of experiments are given directly before and after application as well as at the end of the trial. Statistical analysis was performed at the time-points with the standard deviation (directly before and after application as well as at the end of the trial): (A) After the exposure time of 250 µm glutamate a non-significant reduction of a-wave amplitudes (7.8%) was detected. At the end of the trial, a non-significant reduction of 1.9% was found. (B) Concerning the b-wave, significantly decreased amplitudes of the ERG by 83.7% were recorded. At the end of the trial, a non-significant reduction of 2.3% was noted.
| Time [min] | b-wave amplitude [µV] | SD | a-wave amplitude [µV] | SD |
| 0 | 9.2 | 0.836660027 | -10.4 | 1.140175425 |
| 5 | 9.2 | 1.095445115 | -10 | 1 |
| 10 | 10 | 0 | -10.6 | 0.547722558 |
| 15 | 9.4 | 1.140175425 | -9.6 | 0.547722558 |
| 20 | 9.4 | 0.547722558 | -10 | 1 |
| 25 | 9.4 | 0.894427191 | -10.8 | 0.447213595 |
| 30 | 4.2 | 2.774887385 | -6.6 | 2.50998008 |
| 35 | 4 | 2.449489743 | -5 | 2.236067977 |
| 40 | 3.8 | 1.788854382 | -5 | 1 |
| 45 | 3.6 | 1.341640786 | -5 | 2.121320344 |
| 50 | 2.4 | 1.140175425 | -4 | 1.870828693 |
| 55 | 2.2 | 0.836660027 | -3.4 | 1.140175425 |
| 60 | 2.6 | 0.547722558 | -3.4 | 1.816590212 |
| 65 | 1.8 | 0.836660027 | -2.8 | 1.303840481 |
| 70 | 1.2 | 0.447213595 | -1.4 | 0.894427191 |
| 75 | 4.2 | 0.836660027 | -5.2 | 2.683281573 |
| 80 | 5.8 | 1.303840481 | -6 | 2.828427125 |
| 85 | 6.2 | 1.095445115 | -4.8 | 0.836660027 |
| 90 | 6.8 | 1.095445115 | -6.2 | 2.387467277 |
| 95 | 7.4 | 1.816590212 | -5.6 | 2.302172887 |
| 100 | 6.4 | 0.547722558 | -6.2 | 2.863564213 |
| 105 | 6.6 | 0.894427191 | -7.2 | 2.049390153 |
| 110 | 6.2 | 1.643167673 | -6.4 | 2.966479395 |
| 115 | 8.8 | 3.898717738 | -6 | 3.16227766 |
| 120 | 7.4 | 1.516575089 | -6 | 2.34520788 |
| 125 | 6.8 | 1.095445115 | -6 | 2.645751311 |
| 130 | 7 | 1 | -6.6 | 1.816590212 |
Table 1: Kolmogorov-Smirnov results.
| Time [min] | b-wave amplitude [µV] | SD | a-wave amplitude [µV] | SD |
| 0 | 9.25 | 0.5 | -10 | 1 |
| 5 | 10.5 | 0.577350269 | -10 | 1 |
| 10 | 10.25 | 0.5 | -10.4 | 0.894427191 |
| 15 | 10.5 | 0.577350269 | -9.6 | 0.894427191 |
| 20 | 10 | 0.816496581 | -9.6 | 0.547722558 |
| 25 | 10.75 | 0.5 | -10.2 | 0.836660027 |
| 30 | 3 | 1.825741858 | -8.4 | 2.701851217 |
| 35 | 6 | 3.559026084 | -9 | 1 |
| 40 | 5.25 | 2.62995564 | -8.4 | 2.966479395 |
| 45 | 2.75 | 2.061552813 | -9 | 1.414213562 |
| 50 | 2.75 | 0.957427108 | -8 | 0.707106781 |
| 55 | 2.5 | 1 | -10.2 | 1.095445115 |
| 60 | 2.25 | 0.957427108 | -8.4 | 1.816590212 |
| 65 | 2 | 1.414213562 | -8.6 | 1.140175425 |
| 70 | 1.75 | 0.5 | -9.4 | 2.701851217 |
| 75 | 8 | 3.464101615 | -9.2 | 2.28035085 |
| 80 | 9.5 | 3.696845502 | -9 | 1 |
| 85 | 8 | 2.160246899 | -10.8 | 2.48997992 |
| 90 | 8.25 | 0.957427108 | -9.2 | 2.167948339 |
| 95 | 9.25 | 2.872281323 | -10 | 1.870828693 |
| 100 | 9.75 | 0.957427108 | -9.6 | 1.140175425 |
| 105 | 13.5 | 3.872983346 | -9.2 | 1.643167673 |
| 110 | 11.75 | 2.5 | -9.6 | 1.140175425 |
| 115 | 9 | 1.825741858 | -10 | 1.870828693 |
| 120 | 11 | 3.366501646 | -9.8 | 1.303840481 |
| 125 | 11 | 2.708012802 | -10.8 | 0.447213595 |
| 130 | 10.5 | 0.577350269 | -10 | 1 |
Table 2: Representative measurements for hypoxia for b-wave amplitudes and a-wave amplitudes.
| Glutamate a-wave data | Glutamate b-wave data | Hypoxia a-wave data | Hypoxia b-wave data | |
| D | 0.103827009 | 0.17415038 | 0.195 | 0.125 |
| p-value | 0.928603977 | 0.375501627 | 0.250 | 0.781 |
| alpha | 0.05 | 0.05 | 0.05 | 0.05 |
Table 3: Representative measurements for 250 µM glutamate for b-wave amplitudes and a-wave amplitudes.
