Streptomycetes er jordboende bakterier, der er kendetegnet ved en kompleks udviklingsmæssig cyklus involverer morfologisk differentiering fra en flercellede, næringsstof-latrintømning mycelium til hvilende, unigenomic sporer 1-3.
Under gunstige vækstbetingelser, en typisk Streptomyces Spore begynder at spire ved ekstrudering én eller to mikroberør (figur 1). Disse rør langstrakt med tip udvidelse og vokse ind i en forgrenet hyfe netværk kendt som den vegetative mycelium. Polar vækst og hyfeforgrening er instrueret af den væsentlige protein DivIVA. Denne coiled-coil-protein er en del af en stor cytoplasmatisk kompleks kaldet polarisome, hvilket er afgørende for indsættelse af nye celle kuvert materiale ved strækker spidsen 4-7. Under vegetativ vækst, de svampetrådenes filamenterne bliver ruminddelt af sjældne dannelse af såkaldte cross-vægge 8. Dannelsen af disse tværgående vægge re quires FtsZ, tubulin-lignende cytoskeletale protein der er essentielt for celledeling i de fleste bakterier 9. I Streptomyces, men disse vegetative cross-vægge ikke fører til indsnævring og celle-celle-adskillelse og derfor myceliemassen forbliver som et netværk af indbyrdes forbundne syncytial rum. Som reaktion på næringssaltbegrænsning og andre signaler, der ikke er godt forstået, specialiserede lufthyfer bryde væk fra den vegetative mycelium og vokse i luften 3. Opførelsen af disse strukturer initierer reproduktive udviklingsfase, hvor den lange multi-genomisk lufthyfer blive opdelt i snesevis af lige store unigenomic prespore rum. Denne massive celledeling begivenhed er drevet af den synkrone konstriktion af flere FtsZ ringe inden enkelt sporogenic hyfer 2,10. Morfologisk differentiering er afsluttet ved frigivelse af hvilende, tykvæggede, pigmenterede sporer.
t "fo: holde-together.within-side =" 1 ">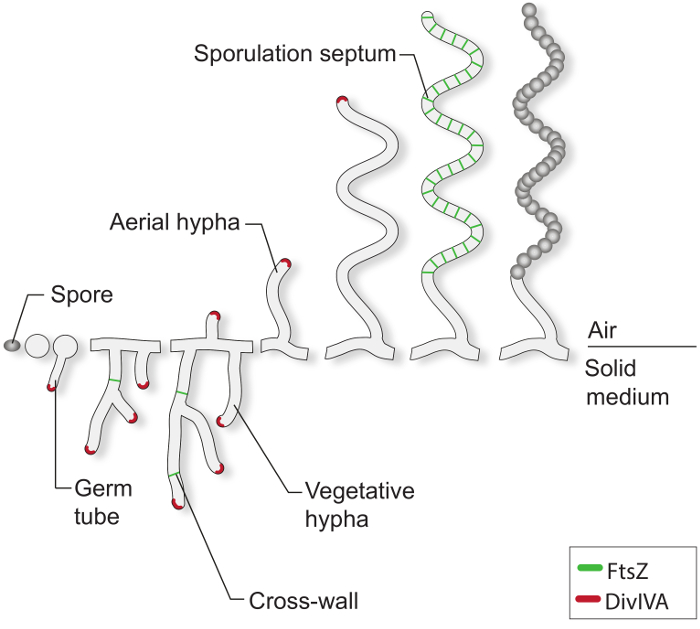
Figur 1:. Den Streptomyces livscyklus på faste medier Dette er en model af livscyklus baseret på klassiske studier af S. coelicolor vokser på agarplader. Den cellulære udvikling af en sporesuspension begynder med dannelsen af en eller to mikroberør, som vokser ved spids forlængelse for at danne et netværk af forgrenende hyfer. Polar vækst og forgrening af den vegetative hyfer er instrueret af DivIVA (rød). Dannelsen af vegetative cross-vægge kræver FtsZ (grøn). Som svar på begrænsninger næringsstoffer og andre signaler, der lufthyfer rejst. Anholdelse af antenne vækst er tæt koordineret med samling af en stige af FtsZ-ringe, som giver anledning til den sporuleringen septa, der inddeler den sporogenic hyfer i box-lignende prespore rum. Disse rum samle en tyk spore- væg og er til sidst Released som modne pigmenterede sporer.
De vigtigste udviklingsmæssige begivenheder i Streptomyces livscyklus er godt karakteriseret 1,3. Men hvad er stadig sparsomme er celle biologiske undersøgelser, der beskæftiger fluorescens time-lapse mikroskopi til at give indsigt i de subcellulære processer understøtter differentiering, såsom protein lokalisering dynamik, kromosom bevægelse og udviklingsmæssigt kontrolleret celledeling. Lev-cell imaging af Streptomyces udvikling har været en udfordring på grund af kompleksiteten af livscyklus og de fysiologiske egenskaber af organismen. Tidligere undersøgelser på den vegetative vækst og de indledende stadier af sporedannelse septumdannelse har ansat oxygenpermeable billeddannende kamre, eller agarose-understøttede vækst af Streptomyces coelicolor på et mikroskop stadium 11-15. Disse metoder er imidlertid begrænset af en række faktorer. Nogle systemer tillader kun kortvarig billeddannelse af cellevækst end fluorescerende proteiner før celler lider under utilstrækkelig ilttilførsel eller vokse ud af fokusplanet på grund af den tredimensionale mønster af hyfe udvikling. I tilfælde, hvor langsigtet billeddannelse er mulig, dyrke celler på agarose puder grænser eksperimentel fleksibilitet, fordi cellerne ikke kan udsættes for alternative vækst- eller stresstilstande, og baggrunden fluorescens fra mediet i agarose puder alvorligt begrænser muligheden for at overvåge svagere fluorescerende signaler.
Her beskriver vi en protokol for live-cell imaging af komplette Streptomyces livscyklus med fremragende præcision og følsomhed. Ved at dyrke Streptomyces i et mikrofluidapparat tilsluttet en fluorescens vidvinklede mikroskop (Figur 2), er vi nu i stand til at overvåge spiring, vegetativ vækst og sporedannelse septumdannelse over en tidsperiode på op til 30 timer. Dette er i høj grad lettes ved anvendelse af de nye model organisme Streptomyces </em> venezuelae fordi det sporulates til nær færdiggørelse i nedsænket kultur og derved overvinder begrænsningen af den klassiske model arter S. coelicolor, som kun sporulates på faste medier 16-20. At hjælpe med at visualisere vegetativ vækst og sporedannelse, vi coudtrykke fluorescens mærkede versioner af cellepolaritet markør DivIVA og nøglen celledeling protein FtsZ.
Vi bruger en kommercielt tilgængelig mikrofluidanordning, der er blevet anvendt med held til mycobakterier, Escherichia coli, Corynebacterium glutamicum, Bacillus subtilis og gær 21-25. Systemet fælder celler i et enkelt fokalplan og tillader kontrol med kontinuerlig perfusion af dyrkningsmedium fra forskellige reservoirer. I detaljeret protokol tager vi fordel af denne funktion til at udsætte S. venezuelae vegetativ mycelium til en ernæringsmæssig nedgearing til fremme sporedannelse.
Protokollen deScribed er for live-cell imaging af hele Streptomyces livscyklus, men alternative medier betingelser eller mikroskop indstillinger kan vælges, hvis specifikke udviklingstrin er af særlig interesse.
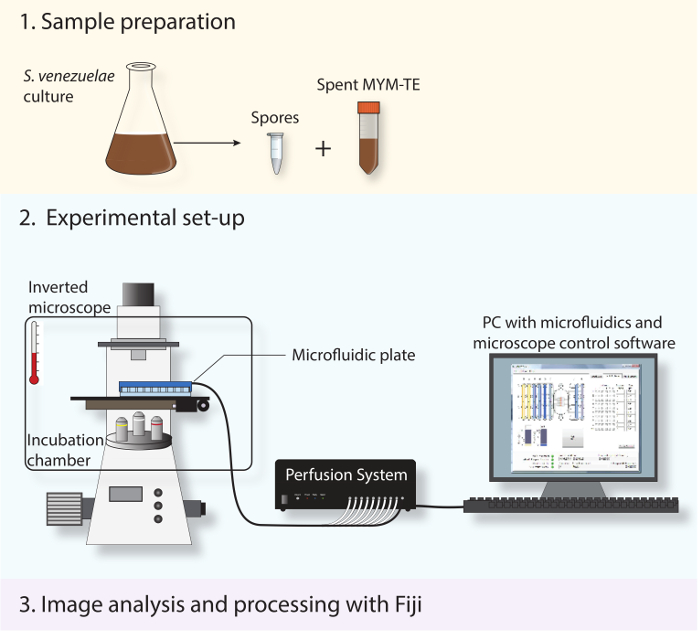
Figur 2: Skematisk afbilder eksperimentelle arbejde-flow. De tre vigtigste trin, der er beskrevet i protokollen, er vist. Først sporer og brugte medium fremstilles ud fra en stationær fase kultur. For det andet er de friske sporer fyldt i en mikrofluidsystem og S. venezuelae afbildes hele dets udviklingsmæssige livscyklus hjælp af et fuldt automatiseret inverteret mikroskop med en inkubation kammer til at opretholde en optimal væksttemperatur. For det tredje er det time-lapse-serien opnåede analyseres og behandles med open source-software Fiji.
