Streptomyces är jordlevande bakterier som kännetecknas av en komplex utvecklingscykel som involverar morfologisk differentiering från en flercellig, närings sophantering mycel vilande, unigenomic sporer 1-3.
Under gynnsamma tillväxtbetingelser, startar en typisk Streptomyces spor att gro genom strängsprutning av en eller två bakterierören (figur 1). Dessa rör avlånga med spets förlängning och växa till en förgrenad hyphal nätverk kallas vegetativt mycel. Polar tillväxt och hyfal förgrening leds av den väsentliga protein DivIVA. Detta coiled-coil-proteinet är en del av en stor cytoplasmisk komplex kallas polarisome, vilket är avgörande för införandet av nya cellhölje material på den utskjutande spetsen 4-7. Under vegetativ tillväxt, de hyfernas trådarna blir fack med ovanliga bildningen av så kallade kors väggar 8. Bildningen av dessa tvärväggar re quires FtsZ, den tubulinliknande cytoskelettala protein som krävs för celldelning i de flesta bakterier 9. I Streptomyces, men dessa vegetativa kors väggar inte leder till sammandragning och cell-cell separation och därför mycelmassan kvarstår som ett nätverk av sammankopplade syncytial fack. Som svar på näringsbegränsning och andra signaler som inte är väl förstådda, specialiserad lufthyfer bryta sig loss från det vegetativa mycelet och växa upp i luften 3. Uppförandet av dessa strukturer initierar den reproduktiva fasen av utveckling, under vilken länge flera genomisk lufthyfer blivit uppdelad i dussintals lika stora unigenomic prespore fack. Denna massiva celldelnings händelse drivs av synkrona sammandragning av flera FtsZ ringar inom enda sporogenic hyfer 2,10. Morfologisk differentiering kompletteras med frisättning av vilande, tjocka väggar, pigmenterade sporer.
t "fo: keep-together.within-page =" 1 ">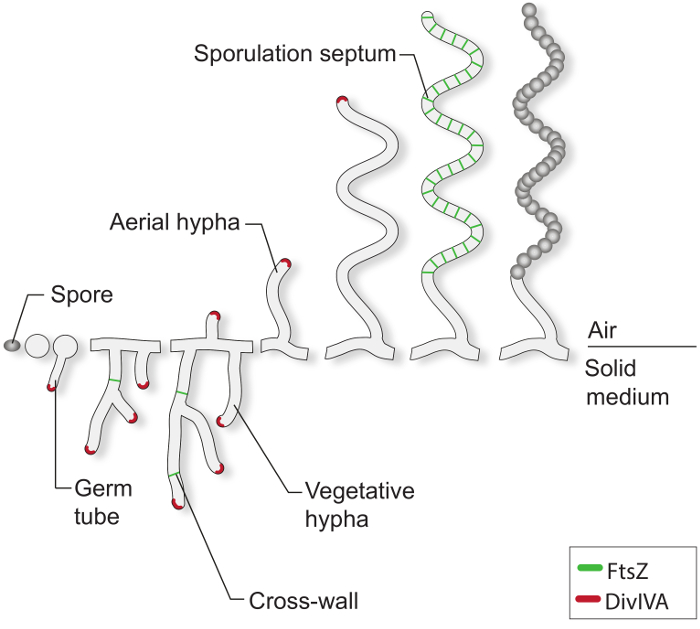
Figur 1:. Streptomyces livscykel på fasta medier Detta är en modell av livscykeln baserad på klassiska studier av S. coelicolor växer på agarplattor. Den cellulära utveckling av en spor börjar med bildningen av en eller två bakterie tuber, som växer genom spets förlängning för att bilda ett nätverk av förgrenande hyfer. Polär tillväxt och förgrening av den vegetativa hyfer riktas av DivIVA (röd). Bildandet av vegetativa tvärväggar kräver FtsZ (grön). Som svar på begränsningar närings och andra signaler är lufthyfer uppfördes. Gripandet av antenn tillväxt tätt samordnas med montering av en stege av FtsZ-ringar, som ger upphov till sporulering septa som compartmentalize den sporogenic hyfer i lådliknande prespore fack. Dessa avdelningar montera en tjock spor vägg och är slutligen Released som mogna pigmente sporer.
De viktigaste utvecklings händelserna i Streptomyces livscykel är väl karakteriserade 1,3. Men vad är fortfarande sällsynta är cellbiologiska studier som använder fluorescenstidsförlopp mikroskopi för att ge insikt i de subcellulära processer som ligger till grund differentiering, såsom protein lokalisering dynamik, kromosom rörelse och utvecklings kontrollerad celldelning. Live-cell imaging av Streptomyces utveckling har varit utmanande på grund av komplexiteten i livscykeln och de fysiologiska egenskaperna hos organismen. Tidigare studier på vegetativ tillväxt och det inledande skedet av sporulering septumbildning har använt syrepermeabla imaging kammare, eller agarosen stödda tillväxten av Streptomyces coelicolor på mikroskop scenen 11-15. Dessa metoder är emellertid begränsad av ett antal faktorer. Vissa system tillåter bara kortsiktiga avbildning av celltillväxt end fluorescerande proteiner före celler lider av otillräcklig syretillförsel eller växa ut ur fokalplanet på grund av den tredimensionella mönster av hyfal utveckling. I de fall där långsiktig avbildning är möjlig, odla cellerna på agaros pads gränser experimentellt flexibilitet eftersom cellerna inte kan utsättas för alternativa tillväxt- eller stressbetingelser, och bakgrundsfluorescens från mediet i agarosen dynor begränsar allvarligt förmågan att övervaka svagare fluorescerande signaler.
Här beskriver vi ett protokoll för levande cell imaging av kompletta Streptomyces livscykel med utmärkt precision och känslighet. Genom att odla Streptomyces i ett mikroflödessystem enhet som är ansluten till en fluorescens widefield mikroskop (Figur 2), kan vi nu att övervaka groning, vegetativ tillväxt och sporbildning septumbildning under en tidsperiod på upp till 30 timmar. Detta underlättas i hög grad genom användning av de nya modellorganism Streptomyces </em> venezuelae eftersom det sporulates till nära färdig i nedsänkt kultur och därmed övervinner begränsningen av den klassiska modellen arten S. coelicolor, som sporulates bara på fasta medier 16-20. För att hjälpa visualisera vegetativ tillväxt och sporulering, vi samar uttrycker fluorescensmärkta versioner av cellens polaritet markören DivIVA och nyckeln celldelning protein FtsZ.
Vi använder en kommersiellt tillgänglig mikroflödessystem enhet som har använts med framgång för mykobakterier, Escherichia coli, Corynebacterium glutamicum, Bacillus subtilis och jäst 21-25. Systemet fångar celler i en enda fokalplan och tillåter kontroll av kontinuerlig perfusion av odlingsmedium från olika reservoarer. I den detaljerade protokollet tar vi fördel av den här funktionen för att exponera S. venezuelae vegetativt mycel till en närings nedväxling för att främja sporbildning.
Protokollet debeskrivs är för levande cell imaging av hela Streptomyces livscykel, men alternativa medier villkor eller mikroskop inställningar kan väljas om specifika utvecklingsstadier är av särskilt intresse.
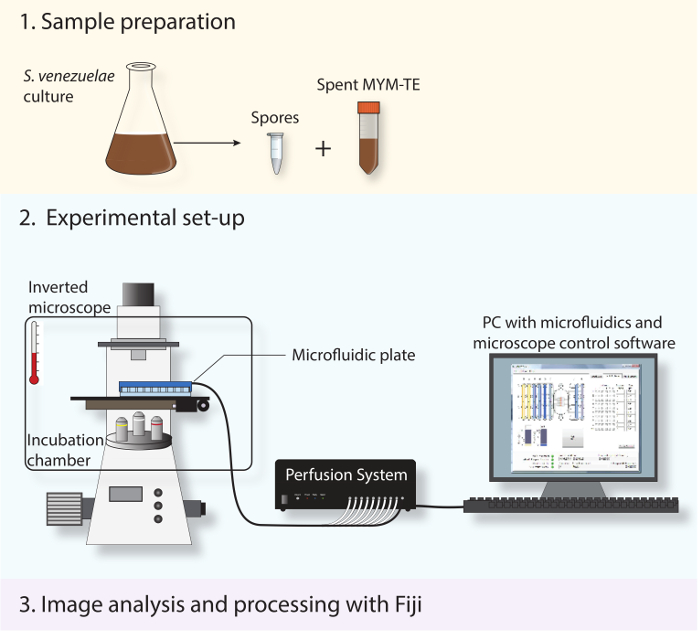
Figur 2: Schematisk skildrar experimentella arbetsflödet. De tre huvudsteg som beskrivs i protokollet visas. Först är sporer och använt medium framställt från en stationär fas kultur. För det andra är de färska sporer laddas in i en mikroflödessystem och S. venezuelae avbildas hela sin utvecklings livscykel med hjälp av en helt automatiserad inverterat mikroskop med en inkubationstid kammare för att upprätthålla en optimal tillväxttemperatur. För det tredje är det tidsförlopp serie erhålls analyseras och bearbetas med hjälp av öppen källkod Fiji.
