Figure 1 shows the process of preparing intact DRG for patch recording. Figure 1A shows the exposure and location of the ganglia after laminectomy. Figure1B shows L3, L4 and L5 DRGs with the nerve roots attached after removing the spinal cord. Then L4 and 5 DRGs are carefully dissected and freed from the vertebrae. Next, the epineurium, a transparent membrane surrounding the DRG, is removed (yellow arrow, Figure 1D). The best location to separate the epineurium is through the site where the dorsal and ventral roots join at the DRG, as indicated by the black arrow in Figure 1D. After peeling off the epineurium, the ventral root is removed and discarded, leaving the dorsal rootlets, spinal nerve and DRG, as shown in Figure 1E. Digestion with collagenase removes the residual epineurium, exposing the neurons and the satellite cells that surround them (Figure 1F).
After completing the DRG preparation, the excitability of small diameter DRG neuron is examined by measuring the rheobase and membrane threshold. As shown in Figure 2A, the rheobase is 260 pA. Figure 2B shows the membrane threshold measured with this method. With the increasing current, the membrane is continuously depolarized until an AP is evoked. In this example the membrane threshold (the potential at which the AP is evoked) is -11.9 mV. Therefore, by maintaining the relationship between the neurons and satellite glial cells, our preparation is closer to the in-vivo situation. Based on our results, the rheobase recorded from small DRG neurons is around 300 pA, and the input resistance is 451.3 ± 27.3 MΩ. Both are higher compared with those measured from dissociated neurons (around 150 pA and 635 MΩ), suggesting that dissociation increased the excitability of DRG neurons 11.
With this preparation, we also measured the inward currents induced by glutamate and allyl isothiocyanate (AITC), which are agonists for glutamate receptors and transient receptor potential cation channel member A1 (TRPA1) respectively. The amplitude of inward currents induced by these ligands will reflect the number of the receptors distributed in the neurons. Figure 3A shows an inward current in a small diameter DRG neuron induced by a 1 sec puff application of glutamate (1 mM). Glutamate induced an inward current (198 pA), which can be largely blocked by co-application of the NMDA receptor antagonist APV and the AMPA/kainate receptor antagonist CNQX (data not shown), confirming the current is mediated by glutamate receptors on the DRG neurons. This data established that there are functional glutamate receptors on DRG neurons. The neuron illustrated in Figure 3B showed inward currents (260 pA) induced by 1 sec puff application of allyl isothiocyanate (AITC, 100 µM), a selective TRPA1 agonist. The inward current was blocked by the selective TRPA1 antagonist 10 µM HC 030031 (data not shown), confirming the currents was mediated by TRPA1 receptors and establishes the presence of TRPA1 receptors on DRG neurons.
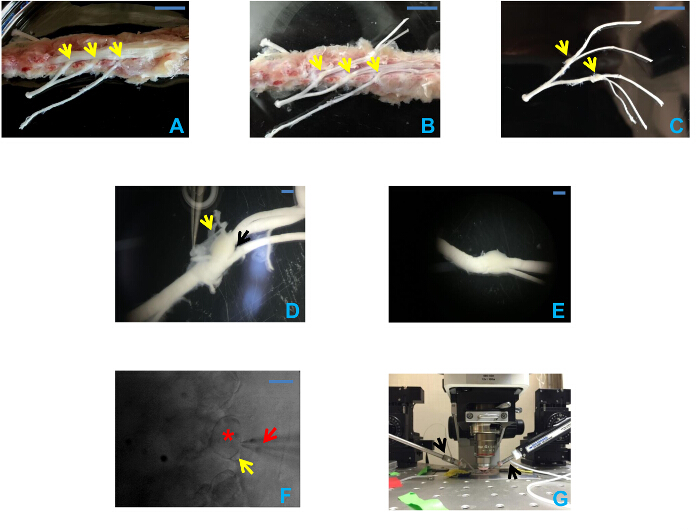
Figure 1: Preparation of intact DRGs. (A) Exposure of the spinal cord after laminectomy and the segmentation of DRGs. Arrows from right to left indicate L3-5 DRGs respectively. Scale bar = 1 cm. (B) Exposure of L3, L4, and L5 DRGs and the attached nerve roots after removal of spinal cord. Arrows indicate L3, L4, and L5 DRGs from right to left. Scale bar = 1 cm. (C) Intact DRGs with nerve roots attached after being dissected from the spinal cord. Arrows indicate L4 and L5 DRGs from right to left. Scale bar = 1 cm. (D) Intact DRG with epineurium attached. In this picture, the epineurium is intact, and the yellow arrow shows the semi-transparent epineurium held by a Dumont forceps. The black arrow indicates the junction formed by dorsal root and ventral root, which serves as a good starting point to peel off the epineurium. Scale bar = 1 mm. (E) The same DRG with the dorsal root attached after the epineurium and ventral root have been removed. Scale bar = 1 mm. (F) Infrared microscope images of DRG following digestion with collagenase. Small- and medium-sized neurons are visible. A satellite glial cell (yellow arrow) is visible surrounding a neuron (asterisk). The red arrow points to the pipette. Scale bar = 10 µm. (G) Recording equipment configuration. The black arrows indicate the pipette holders. The left one holds the drug filled pipette, and recording pipette is on the right. Please click here to view a larger version of this figure.
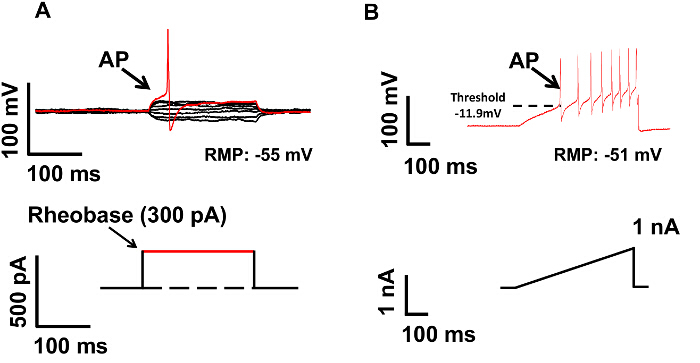
Figure 2: Measurement of Neuronal Excitability in Current Clamp Mode. (A) Rheobase was measured by injecting a series of currents into the DRG neuron (upper panel). The lowest current intensity, which can induce an action potential, is defined as rheobase, as indicated by the arrow in lower panel. The rheobase for this neuron is 300 pA. (B). The membrane threshold was measured by injecting a ramp current. The potential, when an action potential is evoked, is defined as membrane threshold, as labeled by the dashed line in the upper panel. The membrane threshold for this neuron is -11.9 mV. Please click here to view a larger version of this figure.
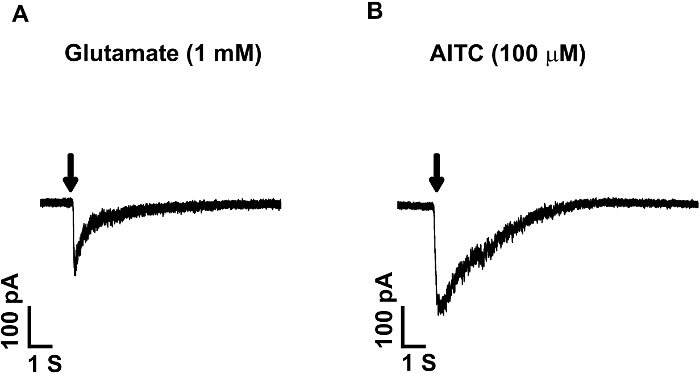
Figure 3: Ligand Induced Currents in Small DRG Neurons. Currents induced by puff application of glutamate (1 mM) or allyl isothiocyanate (TRPA1 agonist, AITC, 100 µM) for 1 sec. Both agonists induced inward currents, suggesting the presence of the glutamate receptors and TRPA1 receptors on small DRG neurons. Glutamate application induced an inward current of 198 pA (A), and AITC induced an inward current of 260 pA (B). Neurons are clamped at -70 mV. Please click here to view a larger version of this figure.
| 2 liters of 10x low-cationic stock | |||
| Final Concentration (mM) | Component | MW | Weight (g) |
| 3 | KCl | 74.6 | |
| 11 | Glucose | 180.2 | |
| 123 | NaCl | 58.4 | |
| 1.25 | NaH2PO4*H2O | 138 | |
| 1 | MgCl2*6H2O | 203.3 | |
| 2 | CaCl2*2H2O | 147 | |
Table 1
| 1 liter of 10x bicarbonate stock | |||
| Final Concentration (mM) | Component | MW | Weight (g) |
| 26 | NaHCO3 | 84.01 | 21.84 |
Table 2
| 100 ml of intracellular solution | |||
| Concentration (mM) | Component | MW | g |
| 130 | KGluconate | 234.24 | |
| 10 | KCl | 74.55 | |
| 10 | HEPES | 238.3 | |
| 10 | EGTA | ||
| 2 | MgCl2*6H2O | 203.3 | |
| Notes: Adjust pH to 7.4 with KOH, and osmolarity to 260 – 280 mOsm with sucrose and distilled water. Add 2 mM MgATP, 0.5 mM Na2GTP and filter before immediate use. | |||
Table 3
