В 1998 году , Огонь и Мелло сообщил , что двухцепочечной РНК (дцРНК) может вызывать ингибирование функции генов в Caenorhabditis Элеганс 1. Эта реакция вызвана дсРНК была названа РНК – интерференция (RNAi), и такие RNAi-опосредованной ген глушителей , как сообщалось, сохраняется у животных, растений и грибов 2-7. У растений и некоторых животных, функции RNAi системно, а это означает , что эффект может распространиться на другие клетки / ткани , где дцРНК не непосредственно вводить (обзор в 8-10). Ученые использовали этот эндогенный клеточный ответ RNAi путем разработки дцРНК целевых генов , представляющих интерес, тем самым сбивая функцию гена без прямого манипулирования геном (обзор в 11-14).
RNAi является мощным инструментом для функциональных исследований из-за следующих преимуществ. Во-первых, даже при минимальной информации о последовательности гена, ген может быть направлена с помощью RNAi. Это особенно важно для стudies не-модельных организмов, не имеющих геномные или транскриптомных данных. Во-вторых, в организмах, где ответ РНКи робастно системным, RNAi-опосредованной нокдаун гена может быть выполнен практически в любом стадии развития. Эта функция очень полезна для изучения функции плейотропных генов. В- третьих, в некоторых случаях, RNAi эффекты распространяются на гонады и потомству, таким образом, что фенотипы наблюдаются у потомства 15,16. Это явление, известное как родительская RNAi (pRNAi), особенно выгодно для генов, влияющих на эмбриональное развитие, а многочисленное потомство производится одним закачиваемой родителем может быть рассмотрено без прямого манипулирования яиц. По этим причинам, pRNAi является методом выбора. Тем не менее, если pRNAi неэффективен, например, для генов, необходимых для оогенеза, то эмбриональная RNAi (eRNAi) должен быть использован. В-четвертых, РНК-интерференция может быть использован для создания эквивалент серии аллельного в том, что количество дцРНК поставляется может варьироваться в диапазоне производить слабого до сильного дефектов, Такая градация фенотипов может быть полезным для понимания функции гена, когда ген участвует в сложном процессе и / или полной потере функции летально. В-пятых, поставка дсРНК как правило, легко и практически осуществимо, особенно у животных, показывая надежные системные реакции RNAi. дцРНК могут быть введены путем микроинъекции 1,5, питание / проглатывания 17,18, замачивание, 19,20 и вирус / доставки бактерии опосредованные 21,22. В-шестых, в отличие от некоторых методов нацеливание / редактирования генов, нет необходимости скрининга для организмов, несущих мутацию или проводить генетические кресты для генерации гомозиготы при использовании RNAi. Таким образом, по сравнению со многими другими методами для изучения функции генов, RNAi быстро, недорого, и может быть применен для крупномасштабных экранов 23-25.
Широкая полезность RNAi обеспечивает средства для проведения функциональных исследований в широком диапазоне организмов, расширение спектра видов для изучения beyond традиционные модельные системы, для которых были разработаны генетические инструменты. Например, исследования с использованием не-модельных систем необходимы , чтобы дать понимание эволюции генов и генных сетей путем сравнения функции ортологах из видов , представляющих различные режимы развития или имеющие различные морфологические особенности 26-29. Эти типы исследований обеспечит лучшее понимание биологического разнообразия, с последствиями как для прикладных и фундаментальных исследований.
Будучи крупнейшей группой животных на планете, насекомые предоставляют прекрасную возможность для изучения механизмов, лежащих разнообразия. Кроме того, насекомые, как правило, маленькие, имеют короткие жизненные циклы, высокая плодовитость, и просты в тыл в лаборатории. За последние два десятилетия, RNAi был успешно применен у насекомых , охватывающих заказы, в том числе двукрылых (мух истинные) 5, чешуекрылые (бабочки и моли) 30, жесткокрылые (жуки) 16,31, перепончатокрылых (sawfложь, осы, муравьи и пчелы) 32, Hemiptera (истинные ошибки), Isoptera (термиты) 34, Blattodea (тараканы) 35, прямокрылые (сверчки, кузнечики, саранча, и katydids) 36 и Phthiraptera (вши) 37. Успешное применение RNAi предоставило функциональные данные для исследований в паттернирования раннего эмбриогенеза (передне-задней оси 32, спинным-вентральной оси 28, сегментация 26,38), определение пола 39,40, хитин / кутикулы биосинтез 41, экдизол сигнализации 42, социальное поведение 43, и многое другое. Методы RNAi , разработанные для разных видов насекомых могут иметь дополнительное преимущество в том , что они, вероятно, будут полезны для борьбы с вредителями (обзор 44-46). RNAi эффекты будут Гено-специфические, а также видовой специфичностью, до тех пор, как несохраняющимся регионы выбраны для таргетинга. Для получения полезных видов насекомых, как пчелами и шелкопрядов, ориентированных на гены, жизненно важных для выживаниявирусы или паразиты , чтобы контролировать инфекцию может обеспечить новую стратегию защиты этих видов 47,48.
Кожеед пятнистый (D. тасиШиз), общее название скрыть жук, распространяется по всему миру , за исключением Антарктиды. Как holometabolous насекомое, жизненный цикл D. тасиШиз включает в себя эмбрионального, личиночного, Куколочные и взрослых стадий (рисунок 1). Потому что он питается плотью, D. тасиШиз используется в музеях для скелетируют мертвых животных и судебно – энтомологи могут использовать его , чтобы оценить время смерти 49,50. D. тасиШиз питается продуктами животного происхождения, включая туш, сушеное мясо, сыр и куколок / коконов других насекомых и тем самым наносит ущерб домашним хозяйствам, которые хранятся продукты питания, и шелк, сыр, и мясной промышленности 51,52. Применение RNAi в этом жука может обеспечить эффективный и экологически чистый способ минимизировать его экономические последствия. Наша лаборатория использовала Д. тасиШиз как новый мОдел насекомых для изучения сегментации 53. Помимо того , что поддается лабораторным разведению, D. тасиШиз представляет интерес для фундаментальных исследований , так как он является разработчиком промежуточного росток, что делает его полезным для изучения видов перехода между кратко- и развития долгосрочных ростка.
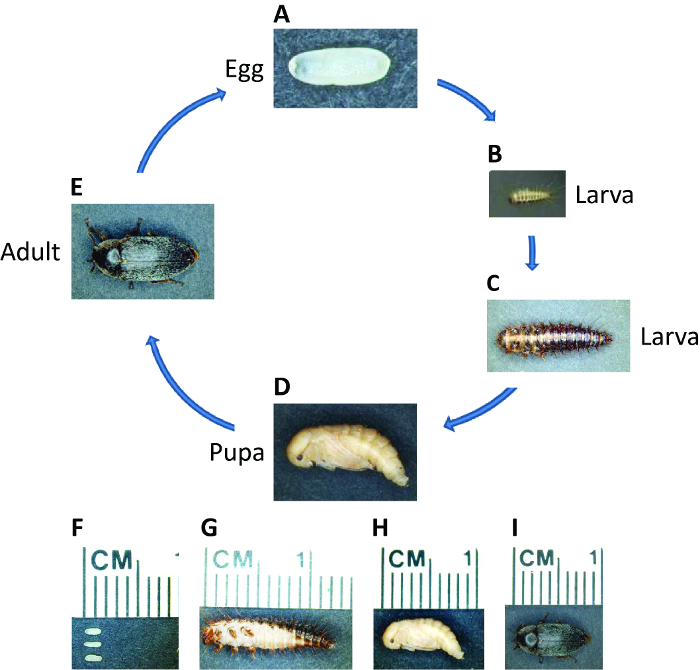
Рисунок 1: Жизненный цикл Д. тасиШиз. Фотографии Д. тасиШиз на разных этапах жизни, как указано. Жизненный цикл от яйца до взрослой особи занимает три недели при 30 ° С, но уже при более низких температурах. (A, F) свежеуложенную эмбрионы от белого до светло – желтого цвета, овальные, около 1,5 мм в длину. Эмибриогенеза ~ 55 часов при 30 ° С. (B, C и G) Личинки имеют темные пигментные полосы и покрыты щетинками. Личинки пройти через несколько возрастах в зависимости от окружающей среды, и их длина может быть увеличен до более чем 1 см. (D, H) </strОнг> Молодые куколки светло-желтые. Окукливание занимает ~ 5 – 7 дней при 30 ° С. (E, I) Вскоре после вылупления, темная пигментация появляется над телом взрослого жука. Взрослые могут жить до нескольких месяцев и одна самка может отложить сотни эмбрионов в течение ее жизни. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
Ранее мы показали , что RNAi является эффективным в нокдаун функции гена в D. тасиШиз 53. Вот наш опыт выращивания D. тасиШиз колонии в лаборатории совместно вместе с протоколами шаг за шагом как для эмбрионального и родительского RNAi настройке, инъекции, уход после инъекции, а также анализ фенотипической. ДцРНК-опосредованные нокдаун и методы анализа ген , введенный здесь не только предоставить подробную информацию для решения вопросов в области D. тасиШиз, но также имеют потенциальное значение Foг применение RNAi в других / видов насекомых, не модель Beetle.
