Male C57bl/6J mice (n=18), aged from 3 to 4 months were included in the study. At day 0 of the experiment, 8 mice underwent a unilateral C4 contusion, right-sided, according to published protocol7,18. As sham procedure, 10 mice underwent a laminectomy on top of C4 without contusion. At day 3, mice were prepared for the intrapleural injections of CTB-fluorophore according to the two different procedures described above. At day 7, all mice were euthanized following anesthesia (ketamine 100 mg/kg and xylazine 5 mg/kg) and exsanguination.
Whole spinal cords were harvested, fixed in paraformaldehyde and cryoprotected in 30% sucrose according to standard lab procedures. Cervical enlargement was isolated, embedded into O.C.T. and cryosectioned longitudinally or transversally at a thickness of 30 µm.
Longitudinal spinal sections were observed with an epifluorescent microscope equipped with filter cube for fluorescence analysis (Excitation: 560/20nm; Emission: 635/30nm). Fluorescent motor neurons in the ventral horn were identified as a linear column of cells from C3 to C5 levels (Figure 6A, upper panel). All labeled cells were located in the ipsilateral gray matter and exhibited morphology consistent with motor neurons (Figure 6B). In injured animals, a striking loss of labeled PhMN was observed at C4 level, together with spinal tissue disruption (asterisk in Figure 6A and 6C). CTB+ PhMNs were manually counted every fifth transverse section (spaced by 150 µm) in uninjured and C4-injured spinal cords (Figure 6C). An estimation of the total number of labeled PhMNs was calculated by multiplying the number of counted CTB+ cells by 5. In uninjured healthy mice, the total number of labeled PhMNs per hemicord was estimated at 151 ± 9 when using transdiaphragmatic (TDia) approach, compared to 178 ± 9 when using transthoracic (TTho) approach (Figure 6D). Following C4 injury, these numbers dropped respectively to 59 ± 14 and 70 ± 10. In this study, no statistical differences were evidenced between the two methods of intrapleural delivery (p>0.05, Mann-Whitney U test; TDia versus TTho).
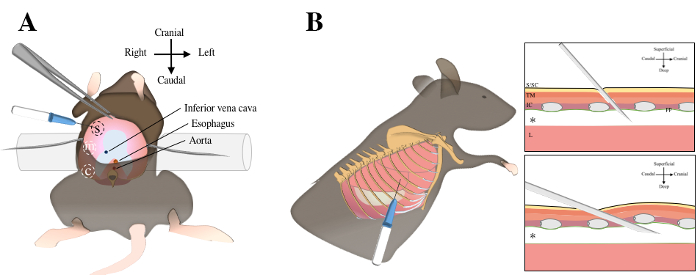
Figure 1. Intrapleural injections using transdiaphragmatic or transthoracic injections. Transdiaphragmatic approach includes a laparotomy, exposure of abdominal surface of the diaphragm (in pink color) and three injections in the sternal, medial and crural regions A. For transthoracic approach, the mouse is laid in left lateral decubitus position. The sixth and seventh ribs are identified under the elbow region by manual palpation B. The needle is inserted through the skin, the subcutaneous tissue, the thoracic muscles, the intercostal muscles and the parietal pleura to reach the pleural cavity (asterix). S, sternal; m, medial; c, crural; S/SC, skin and subcutaneous tissue; TM, thoracic muscles; IC, intercostal muscles; pp; parietal pleura; L, lung. Please click here to view a larger version of this figure.
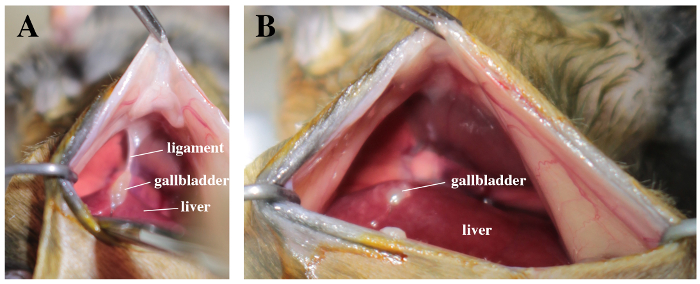
Figure 2. Surgical field with a view on liver, gallbladder and abdominal face of the diaphragm. A. Note the suspensory ligament between the gallbladder and the diaphragm. B. The ligament is carefully cut off. Please click here to view a larger version of this figure.
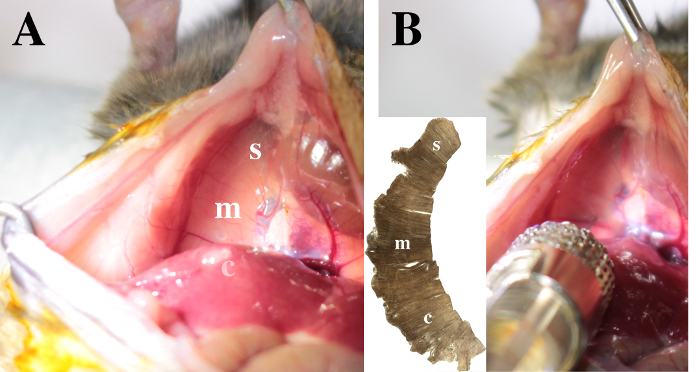
Figure 3. View on the diaphragm. The sternal (s), medial (m) and crural (c) areas are identified in the right hemidiaphragm. A. Note that the crural region is posterior to the liver lobes. B. To inject CTB in the pleural space, insert the needle 1 mm deep through the right hemi-diaphragm. The injection in the medial area is illustrated. Please click here to view a larger version of this figure.
Figure 4. Transthoracic approach. Mouse is positioned in left lateral decubitus A. Identify the costal margin of the diaphragm and the elbow region. B. The sixth and seventh intercostal spaces are identified deep to the elbow region. Please click here to view a larger version of this figure.
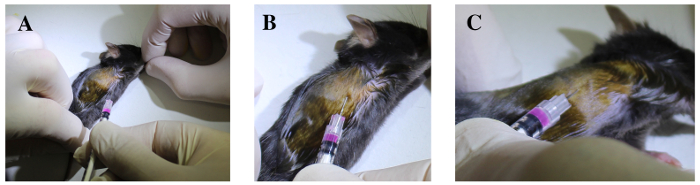
Figure 5. The sixth and seventh intercostal spaces are identified deep to the elbow region. A. Fore- and hind- limbs are extended. B. The syringe is cranially oriented and tangentially inserted under the sixth or the seventh rib, 3 mm deep through the intercostal muscles, with the bevel upside-down. C. The needle is gently elevated and the ribs are lifted to confirm the needle is well positioned into the thoracic cavity. Please click here to view a larger version of this figure.
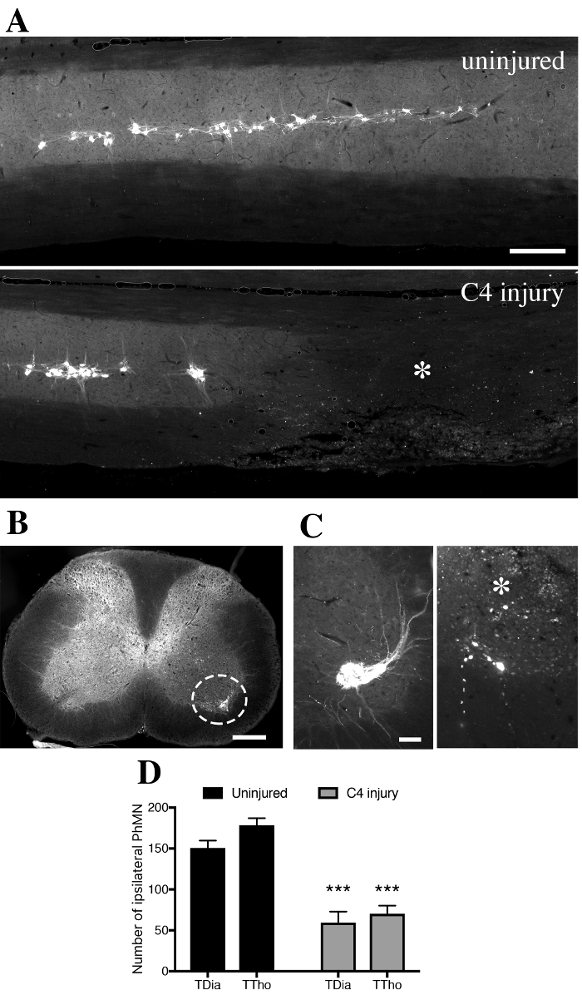
Figure 6. CTB+ cells in the ventral horn gray matter of intrapleurally-injected mice. A. In spinally-uninjured mouse, CTB+ phrenic motor neurons distribute along the cervical spinal cord from level C3 to C5. In contused mouse, tissue disruption is observed unilaterally with loss of CTB+ phrenic motor neurons at C4 level (asterix). B. Labeled CTB+ cells were located in the ipsilateral gray matter and exhibited morphology consistent with motor neurons. C. Transverse sections of spinal cords were used to quantify the number of CTB+ cells at specific distances along the spinal cord in uninjured (left) and injured (right) mice. D. Quantification of CTB+ cells found in the ipsilateral hemicord in uninjured or injured mice, according to the transdiaphragmatic (TDia) or transthoracic (TTho) approach of intrapleural CTB delivery. Bars represent 250 µm. Data were expressed as the mean ± standard error of the mean (SEM). n= 5-7 mice in each TDia group; n= 3 mice in each TTho group. Non-parametrical Mann-Whitney U tests were performed and results were considered as significantly different for p<0.05. ***p<0.001 for uninjured versus C4 injury. Please click here to view a larger version of this figure.
