When this protocol is performed properly, one can determine the effects of aged and/or diseased human Bruch's membrane on RPE cell function by the establishment of polarity and the outer blood retinal barrier, polarized secretion of growth factors and cytokines, and phagocytosis of photoreceptor rod outer segments.
We have generated data that demonstrate a disease phenotype on aged human Bruch's membrane. For example, culturing RPE cells on aged human Bruch's membrane explants decreases RPE cell reattachment (Figure 3). RPE cells cultured on aged human Bruch's membrane decreases the capacity of these cells to phagocytize rod outer segments (ROS), a critical RPE function (Figure 4). Moreover, apoptotic cells can be identified and compared on these button explants using a terminal deoxynucleotidyl transferase dUTP nick end-labeling (TUNEL) stain as described in previous work26. The effects of an aged and/or diseased human Bruch's membrane on RPE cell gene expression profile can be determined using microarray technology. We have demonstrated that RPE cell gene expression is altered when cultured on human Bruch's membrane from older individuals compared to explants from younger individuals (Figure 5)27. For statistical analysis, explants from at least 3 – 5 human donors are used per group.
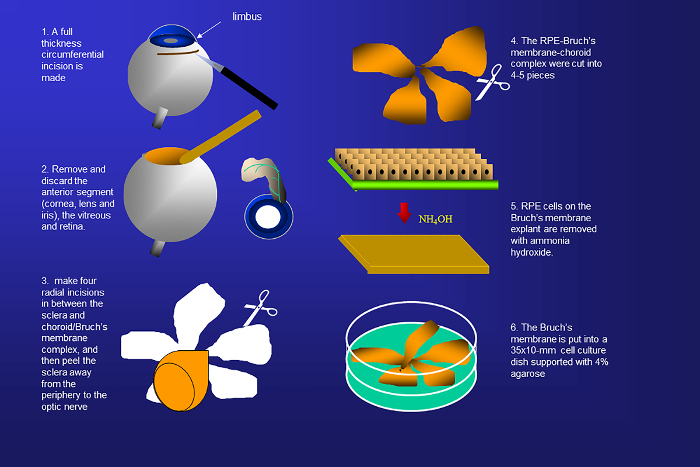
Figure 1. Schematic representation of the isolation of human Bruch's membrane from donor eyes. Human Bruch's membranes are harvested by removing the sclera, anterior segment, retina, vitreous, and native retinal pigment epithelial (RPE) cells. Please click here to view a larger version of this figure.
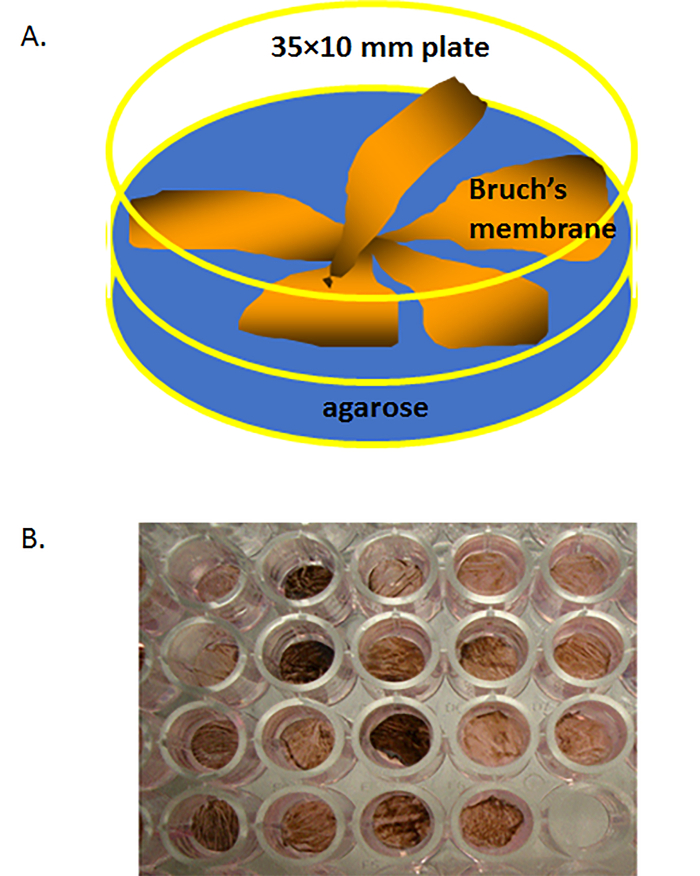
Figure 2. Isolated human Bruch's membrane (BM) in a culture dish. (A) Schematic of human Bruch's membrane explants (each 1.5 inches in diameter, with solidified agarose gel) placed in a 60 × 15 mm polystyrene cell culture dish (BM face-up) filled with warm liquid agarose on the bottom of dish. (B) A human Bruch's membrane explant that has been trephined into a few 6 mm circular buttons (explants), each ow which was then placed on 4% agarose at 37 °C in a non-treated polystyrene well of a 96-well plate. Please click here to view a larger version of this figure.
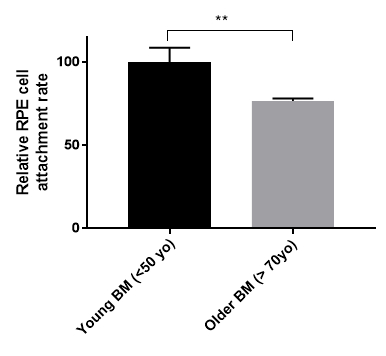
Figure 3. Retinal pigment epithelial (RPE) cell reattachment rate on young and older human Bruch's membrane (BM). Primary RPE cells were seeded onto human Bruch's membrane explants from young (<50-year-old, n = 3) or older (>70-year-old, n = 5) donor eyes for three weeks. Cell attachment rate was measured by MTT cell viability assay. Older human Bruch's membranes reduced the RPE cell attachment rate by 24% (young BM 100 +/-8.54 S.E. vs. older BM 76.65 +/-1.44 S.E.). **p <0.05, S.E. = standard error.
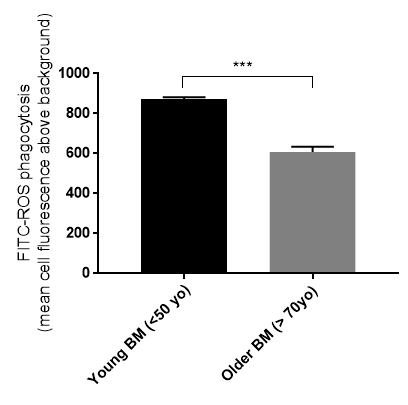
Figure 4. Retinal pigment epithelial (RPE) cell phagocytosis is affected by the age of the human Bruch's membrane (BM). Primary RPE cells cultured on older human Bruch's membrane explants had a reduced ability to phagocytize rod outer segments (ROS) (young BM 873 +/-9.3 S.E., n = 5 vs. older BM 608 +/-25.4 S.E., n = 5). ***p <0.01, S.E. = standard error.
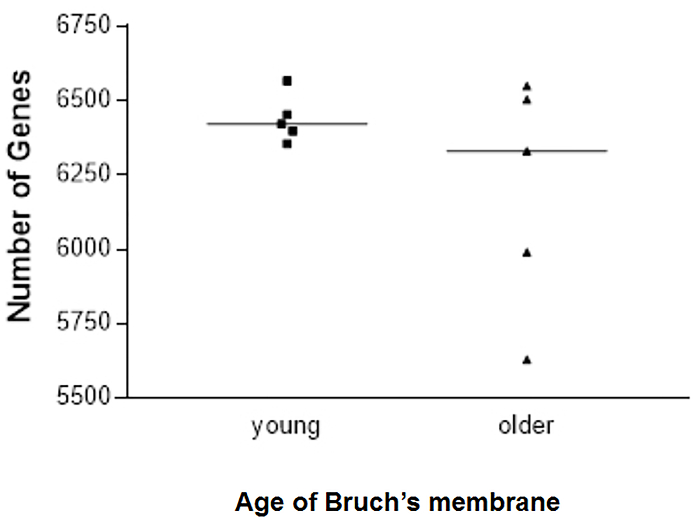
Figure 5. Number of retinal pigment epithelial (RPE) cell genes expressed in each sample cultured on young or older human Bruch's membrane explants. There was more scatter in the number of genes expressed in primary RPE cells seeded onto older versus younger human Bruch's membrane (6201 ± 388 vs. 6439 ± 80, respectively); five explants were tested for each age group. Presented with the full permission of all authors of Cai, H et al.27. Please click here to view a larger version of this figure.
