Kræft er en af de førende dødsårsager i verden. Derfor er der et krav om at udvikle pålidelige eksperimentelle sygdomsmodeller for at opnå en bedre forståelse af sygdommen samt for at udforske potentielle terapeutiske tilgange. En af de mest almindeligt anvendte eksperimentelle in vivo modeller til at studere hudkræft udvikling er den kemisk induceret to-trins hud carcinogenese model1,2. Modellen giver et værktøj til at studere tumor initiering, forfremmelse, og progression i tillæg til specifikke begivenheder såsom immuncelle infiltration og angiogenese.
At bruge to-trins hud carcinogenese model, bagsiden hud af mus er behandlet med to forskellige kemikalier, der tilsammen fremkalde tumordannelse. Modellen initieres med en lav dosis af mutagen, DMBA, efterfulgt af længerevarende udsættelse for tumor promotoren, TPA3 (figur 1). Dmba muserer DNA tilfældigt ved at danne kovalente addukter med DNA fra Epidermal celler og primære keratinocyt stamceller4,5,6,7. Nogle af disse tilfældige mutationer finder sted i en proto-oncogene, såsom Hras1 (mutationer i Kras og NTM opdages også) og omdannelsen af proto-onkogener til onkogener driver tumordannelse under ordentlige stimuli. TPA, igen, er den mest almindeligt anvendte tumorvækst fremmende middel. Dens molekylære mål er proteinkinase C (PKC)8. TPA aktiverer også WNT/β-køreledningen i signalering, der er afgørende for tumordannelse i model9. Gentagen og langvarig udsættelse for fremme agent fører til øget celle signalering, øget produktion af vækstfaktorer, og en lokal inflammatorisk reaktion, som er indlysende på grund af øget DNA-syntese og inflammatorisk celleinfiltration i den behandlede hud.
De vigtigste inflammatoriske mediatorer i DMBA-TPA-modellen er blevet identificeret10. Interleukin-17a (Il-17a) er kendt for at være særligt tumorigent i dmba-TPA-modellen11,12. Det virker i synergi med interleukin 6 (Il-6) og deltager i makrofag og neutrofilrekruttering13,14. Desuden har CD4+ T- celler og neutrofiler vist sig at være tumorigent i dmba-TPA-modellen. Endelig kan makrofager også fremme tumor i model15,16,17.
Under forfremmelses fasen forstærkes celle spredningen af de muterede celler, og et vedvarende hyperplasi af epidermis vedligeholdes1. Dette fører til papilloma udvikling i huden i 10 – 20 uger, hvorefter papillomer begynder at konvertere til maligne tumorer, plade celle karcinomer (SCCS)2. Men mindre end 10% af papillomer fremskridt til malignitet, selv om denne procentdel også afhænger af den genetiske baggrund af musene2,18. I årtier var det ikke kendt, hvilken type celler blev oprindeligt muteret i tumorer fører til malignitet, selv om nogle undersøgelser havde rapporteret klart forskellige funktioner i de Maligne tumorer i forhold til benign papillomer19,20. Men, nylige undersøgelser har i høj grad øget vores forståelse på den klonale oprindelse af tumordannelse i DMBA-TPA model21. 22. 23. det blev påvist, at både knoglemarv-afledte epiteliale celler og hårsæk stamceller bidrager til tumordannelse22. Stage-specifikke Lineage Tracing undersøgelser har afsløret, at godartede papillomer af monoklonale oprindelse, men de rekruttere nye epiteliale cellepopulationer21,23. Men kun en af celle klonerne fungerer som en drivkraft for carcinogenese; den indeholder en HRAS-mutation23. Progression til karcinom dannelse er forbundet med en klonal feje23.
Det kræftfremkaldende DMBA initierer papilloma dannelse og TPA fremmer tumorvækst. Derfor kan tumor initiering undersøgt separat fra kampagnen ved at afbryde eksperimentet før TPA-behandlingsperioden. Som tumorprogression er undersøgt ugentligt det giver en stor mulighed for detaljeret tumorvækst analyse i hele studiet. Fordi tumorer er genereret af eksterne kemikalier, en onkogen mutation i kimcelle er unødvendig. Således er det ligetil at studere virkningerne af en genetisk baggrund (f. eks. knockout/transgene vs. vildtype) på tumor2. I sum, DMBA/TPA Skin Cancer model er en særlig nyttig tilgang til at studere rollen af immunsystemet i tumorprogression samt til evaluering af tumor initiering og forfremmelse skridt selvstændigt eller indbyrdes afhængige.
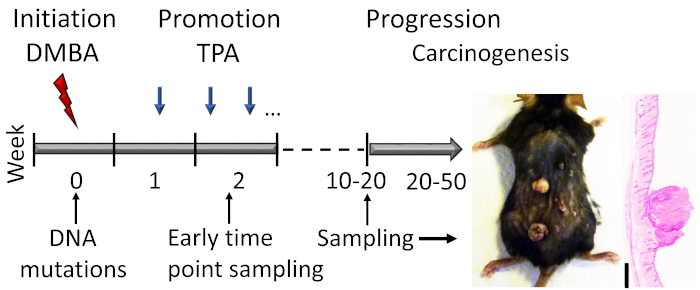
Figur 1: DMBA-TPA-induceret hud carcinogenese model skitse. Det kræftfremkaldende dmba påføres topisk for at inducere DNA-mutationer i Initierings fasen af modellen. Den vækstfremmende agent TPA administreres 2x om ugen for at øge celle spredningen under forfremmelses fasen, hvilket fører til udviklingen af papillomer i huden. Dyr ofres, efter at papillomas respons når et plateau, sædvanligvis inden for uger 15-20, afhængigt af muternes genetiske baggrund. En lille del af papillomer kan videreudvikle sig til VKF inden for 20 – 50 uger. For at studere tidlige hændelser i Initierings fasen og den tidlige forfremmelses fase kan der indsamles prøver (f. eks. kort tid efter den anden TPA-applikation). Et repræsentativt fotografi og hematoxylinlegemer og eosin farvede tværsnit af papillomer på en C57BL/6 mus hud efter 19 ugers behandling er vist. Scale bar = 0,1 mm. Klik her for at se en større version af dette tal.
