Prosedyren for kromatisk korreksjon er en avveining mellom nøyaktighet og innsats. For å spare unødvendig innsats, er det bedre å vite hvor mye nøyaktighet som kreves for studien din. Den høyeste nøyaktigheten er kanskje ikke nødvendig for konvensjonell bredfelt (live) bildebehandling, og dermed er lyse feltreferansebilder ofte tilstrekkelige til å korrigere kromatisk skift. På samme måte, når bildetilstanden og miljøet er konstant, vil gjentatt bruk av en biologisk kalibrering spare tid. På den annen side, hvis en svært nøyaktig registrering er ønsket, er det nødvendig med krysstale av høy kvalitet eller biologiske kalibreringsreferansebilder. For best mulig ytelse bør referansebilder oppnås med så like vilkår og tidspunkter som målbildene som mulig. Så lenge både referanse- og målbilder oppnås av samme mikroskopi, vil høyere romlig oppløsning forbedre korrigeringsnøyaktigheten. Hvis dekonvolutsjon er tilgjengelig for både referanse- og målbilder, kan implementering av dette før korrigering forbedre korrigeringsnøyaktigheten. For best mulig ytelse bør prøvetakingsteoremet for den optiske aksen (Z) oppfylles i både referanse- og målfilen for nøyaktig subpixel interpolering (protokolltrinn 2.1.3).
Unnlatelse av å korrigere kromatisk skift fører til feil konklusjoner. Videre kan bruk av feil kalibrering til og med forverre kromatiske skift i stedet for å korrigere dem, og dette må derfor unngås. Vi har oppsummert mulige årsaker til feil, og deres felles løsninger, i tabell 2. For å undersøke årsaken til en feil, er det i første omgang nødvendig å visuelt kontrollere om det kromatiske skiftet i referansebildet er nøyaktig korrigert (protokolltrinn 3.12). De fleste feil skyldes kvaliteten på referansebildene og er lett utbedret i henhold til beskrivelsene i tabell 2. Når det gjelder kvaliteten på referansebilder, er det viktig å merke seg at nøyaktigheten av global justering reduseres hvis hele synsfeltet ikke er fylt med prøven (Figur 7, Tabell 2). Sammenlignet med det gode eksemplet som vises i figur 7A,inneholder det dårlige eksempelet vist i figur 7B bare tre kjernefysiske konvolutter i øvre venstre region, og Chromagnon klarte ikke å justere en del av dette bildet. Dette er fordi den globale justeringsmetoden for Chromagnon deler synsfeltet i fire regioner (Figur 7C) for å måle forskjellene i rotasjon og forstørrelse med høy nøyaktighet3. Denne metoden, hvis den er riktig drevet, er en ordre mer nøyaktig enn andre lineære metoder som loggpolpolering og simplex-metoder3. Hvis noen av de fire regionene ikke er tilgjengelige, vil Chromagnon bytte til mindre effektive lineære metoder. Derfor, for best ytelse, eksemplene vist i figur 7B og figur 7C er uønsket, og de fire regionene skal fylles med objekter. Brukere kan sjekke om et kvadratisk område i synsfeltet ikke er tilgjengelig for måling ved å se på loggfilen (“Chromagnon.log”; se protokolltrinn 3.10). Heldigvis kan dette problemet lett overvinnes ved å snitte flere biologiske kalibreringsbilder eller bruke lokal justering for krysstale- eller lysfeltreferansebilder (tabell 2). I motsetning til tilfelle av unnlatelse av å korrigere referansebilder, er det vanskeligere å identifisere å ikke korrigere målbilder. Fordi slike feil oppstår på grunn av forskjeller i filformater, bildebehandlingsforhold, bildebehandlingstid, bildebehandlings-/justeringsmetoder mellom referanse- og målbildene(tabell 2),bør brukerne alltid være forsiktige når de bruker referansebilder som er oppnådd under forskjellige forhold/tidspunkter fra målbildene. Noen eksempelbilder er tilgjengelige for testing (https://github.com/macronucleus/Chromagnon) for å få konkrete ide om de gode og dårlige eksempelbildene.
| Problem | Forårsake | Løsning |
| Kan ikke rette opp referansebildet | Lav kontrast | Få et høyere kontrastbilde hvis mulig. Hvis et referansebilde for lysfelt brukes, må du hente bildet i en vannbasert løsning for å oppnå høyere kontrast av cellen. Alternativt kan du prøve å bruke beregningsstøyreduksjon (f.eks. gaussisk filtrering). Slå av lokal justering, som er mer følsom for støy. |
| Kontaminering av ikke-relaterte bilder | Fjern kilden til de ikke-relaterte bildene i eksemplet hvis mulig. For krysstalereferansebilder, sjekk eksitasjonsspektraen på fargestoffene som brukes til målbildene. Hvis fargestoffene er begeistret under oppkjøpet av et crosstalk-bilde (f.eks. Alexa Fluor 568 eller 594), bør du vurdere andre fargestoffer (f.eks. Alexa Fluor 555). Hvis støv på kamerabrikken skaper en åpenbar kanalforskjell, rengjør du kamerabrikken eller bruker en beregningsmessig flatfeltmetode. | |
| Et ekstremt lyspunkt laget av en kosmisk stråle | Få bildet igjen hvis mulig. Alternativt kan du prøve å bruke beregningsstøyreduksjon (f.eks. median eller gaussisk filtrering). | |
| Dekonvolutasjonsartefakter (kunstige signaler på aksiale og laterale kanter) | Trim kantpikslene eller Z-delene etter dekonvolusjon. Hvis den ene siden er trimmet, bør den andre siden også trimmes for å opprettholde bildesenteret. | |
| Z-trinnstørrelsen er for sparsomme | En Z-stabel bør anskaffes for å oppfylle Nyquist-kriteriet som skrevet i protokoll 2.1.3. | |
| Optisk avvik | Sfærisk avvik er det store avviket forårsaket av brukere. Velg riktig objektiv for prøven og bruk en dekksliptykkelse på 170 μm. Hvis objektivet er utstyrt med en korreksjonsring, justerer du den for å finne posisjonen der det høyeste fluorescensantallet er hentet fra fokuset. Når det gjelder et oljenedsenkingsmål uten korreksjonsring, juster brytningsindeksen for nedsenkingsoljen som øker fluorescenstellingen i fokus. | |
| Synsfeltet er ikke fylt opp (fig. 7) | Når det gjelder biologiske kalibreringsreferansebilder, gjennomsnittlig mange bilder. I tilfelle av krysstale eller lysfeltreferansebilder, bruk lokal justering. | |
| En uidentifisert programvarefeil | Rapporter problemet via GitHub (https://github.com/macronucleus/Chromagnon/issues) | |
| Kunne ikke rette målbildet | Metadata for bildefilen går tapt | Bruk det opprinnelige mikroskopfilformatet som inneholder fullstendige metadata, og unngå å konvertere til en tiff-fil med flere sider før behandling. Bruk samme rekkefølge av kanaler som skrevet i protokoll 3.3. |
| Feil justeringsmetoder for den gitte mikroskopien | Ikke bruk den lokale justeringsmetoden når du måler fra biologiske kalibreringsreferansebilder til målbilder. Ikke bruk andre krysstalereferansebilder enn wide-field mikroskopi. | |
| Forskjeller i bildebehandlingsforhold | Hold bildeforholdene konstant mellom referansen og målbildene som skrevet i protokoll 2.3.3. | |
| Forskjeller i prøve (inkludert coverslip) | Bruk alltid samme monteringsmedium, dekkslip (f.eks. 1,5H) og en lignende fokusdybde. | |
| Mikroskopdrift siden kalibreringen sist ble gjort | Lag en kalibrering så ofte som annenhver uke. Hold temperaturen konstant, og bruk et flytende bord for å unngå maskinvaredrift av mikroskopet. |
Tabell 2: Feilsøking for kromatisk korrigering.
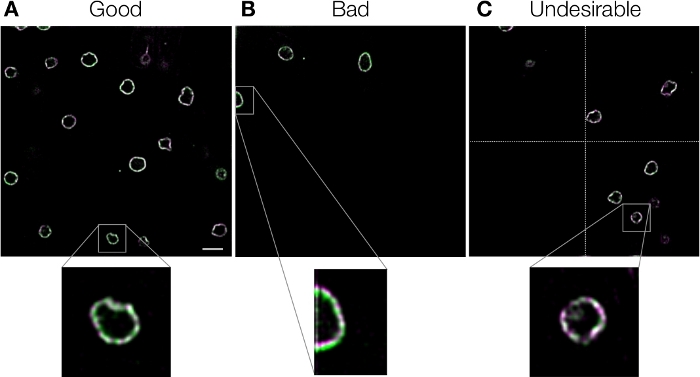
Figur 7: Eksempler på referansebilder. Kjernefysisk konvolutt i fisjon gjær celler merket med GFP og mCherry. Bilder ble anskaffet med konvensjonell bredfelts mikroskopi. Kromatiske skift ble korrigert ved hjelp av Chromagnon uten lokal justering ved hjelp av bildene selv som referansebilder. Bilder ble deretter dekonvolvert for å vise detaljene. (A)Et godt eksempel med mange objekter i synsfeltet. (B)Et dårlig eksempel med objekter bare øverst til venstre. Feiljustering er åpenbar på en bestemt region av bildet. (C)Et uønsket eksempel der en av kvarsjonen (atskilt med stiplede krysslinjer) er tomt. Skalalinje i panel A indikerer 5 μm for full feltvisning og 1,25 μm for forstørret visning og gjelder for alle paneler. Vennligst klikk her for å se en større versjon av dette tallet.
I denne protokollen beskrev vi tre forskjellige referansetyper (tabell 1). Blant dem trenger krysstale referansebilder og biologiske kalibreringsreferansebilder ytterligere nøye diskusjon. For krysstalereferansebilder kan prøver farget med DAPI eller Hoechst 33342, og montert i glyserol eller kommersielle monteringsmedier effektivt brukes til å justere de blå, grønne og røde kanalene. På samme måte kan Alexa Fluor 488 brukes til å justere de grønne og røde kanalene. Men å skaffe crosstalk fluorescens er ofte vanskelig siden mange blå fargestoffer unntatt DAPI og Hoechst er dimmer og forfall raskere enn de fleste grønne og røde fargestoffer. Videre er utslippsspektraen av moderne fargestoffer smalere, noe som gjør justeringen av mer enn tre kanaler ved denne metoden utfordrende. Det bør også tas hensyn til noen vanlige røde fargestoffer (f.eks. Alexa Flour 568 og 594, men ikke Alexa Fluor 555) som kan være begeistret av fiolett lys, som forhindrer å få høykontrast krysstalebilder fra blå fargestoffer. En annen ulempe er at denne metoden ikke kan måle kromatisk aberrasjon av eksitasjon lysbaner i flerfarget eksitasjon, fordi bare en enkelt eksitasjon bølgelengde brukes til eksitasjon (Tabell 1). Som de fleste avanserte mikroskopi bruker endret belysning optikk, er anvendelsen av denne metoden begrenset. Likevel er den høyere korreksjonsnøyaktigheten tilstrekkelig fordelaktig for at den skal beskrives i denne protokollen. Generelt bør et krysstalebilde tas etter et målbilde for å forhindre bleking eller fototoksiske effekter. For SMLM observert med bredfeltsmodus, bør et referansebilde anskaffes før du anskaffer et målbilde, da fluorescensfarger kan blekes mens bildebehandling.
Biologiske kalibreringsreferansebilder gjør det enkelt for brukere å justere et ønsket antall kanaler på bekostning av ekstra prøveklargjøring. En annen fordel med biologiske kalibreringsreferansebilder er tilgjengeligheten av “snitt” flere referanser som bidrar til å fylle alle synsfelt. Denne metoden kan lide av forskjeller i bildeforhold hvis kalibreringsprøven er klargjort på et annet lysbilde. Det meste av dette problemet kan løses hvis både mål og referanser er utarbeidet på samme lysbilde ved hjelp av kommersielle kamret coverbriller (Tabell 1), og andre bildebehandlingsforhold holdes konstant som i protokolltrinn 2.3.3. I dette tilfellet kan en korrigeringsnøyaktighet som ligner på krysstalereferansebilder forventes3. Protokollen for å bruke phalloidin som vist her er en av de enkleste måtene å flekke en enkelt cellulær struktur med flere farger. Det er mange mulige scenarier for å forberede biologiske kalibreringsprøver. For immunstoppnåelse kan en prøve merkes med et enkelt primært antistoff etterfulgt av farging med sekundære antistoffer i flere farger. På denne måten kan en enkelt målstruktur merkes med flere farger. Alternativt, 5-ethynyl-2′-deoxyuridine, oppdaget av “klikk” kjemi etiketter nylig syntetisert DNA i flere farger med høy tetthet, som beskrevet i detalj tidligere8. For levende celler er det nyttig å forberede en transgen stamme som skjuler to kopier av et gen som er smeltet sammen til GFP eller mCherry for å merke samme struktur med to farger. Hvis kopinummeret til genet er kritisk som ofte observert for membranproteiner, kan en enkelt kopi av genet smeltes sammen til GFP og mCherry (figur 7). Fotokonverterbare fluorescerende proteiner, som mEOS218,kan også brukes ved å belyse et moderat nivå av fiolett lys for å oppnå både proteinarter med eller uten fotokonvertering. Under lave oksygenforhold kan GFP også brukes som et fotokonverterbart protein fra grønt til rødt19,,20. Hvis du velger riktig kalibreringsprøve, blir eksperimentet mer robust.
