Drevet av den økende anvendelsen av neste generasjons sekvenseringsteknikker, kan romlig oppløsning i utviklingsbiologi økes. Moderne studier tar sikte på å dissekere vev ned til spesialiserte celletyper, om ikke encellede nivå1,2,3,4. For dette formål har en mengde forskjellige metoder blitt utarbeidet i løpet av de siste femti årene (se figur 1A)5,6,7,8,9,10,11,12,13,14,15.
Mange verktøy i plantevitenskap har vært tilpasninger av teknikker som var pionerer i dyreforskning. Dette er ikke tilfelle for metoden vi introduserer i detalj her. I 2005, utstyrt med sterk bakgrunn i proteinoversettelse, satte Bailey-Serres Lab ut for å konstruere ribosomale proteiner for påfølgende affinitetsrensing16. Dermed kunne de unngå tidkrevende og arbeidskrevende polysom profilering, som er basert på ultracentrifugering med en sukrosegradient og ble brukt til å vurdere å oversette ribosomer siden 1960-tallet17,18. Metoden har siden blitt referert til som translasjonell ribosom affinitet rensing (TRAP)16. Etter vellykkede translatome studier i planter, Heiman et al. tilpasset TRAP for dyr19 og andre utvidet sin søknad til gjær20, Drosophila21, Xenopus22 og sebrafisk23,24.
Selv om genetisk modifisering av modellsystemet er en forutsetning for TRAP, som begrenser anvendelsen til arter som er egnet til genetisk transformasjon, kan man samtidig utnytte denne innvendingen mot undergrupper av celler som er av spesiell interesse og ellers ekstremt vanskelig å isolere fra det intakte vevet/orgelet25 (f.eks. svært forgrenede dendrittiske celler i en mushjerne eller sopphyphae i infisert plantevev). I planter holdes alle celler på plass via cellevegger som danner grunnlaget for det hydrostatiske skjelettet26. For å frigjøre en plantecelle fra denne matrisen, har forskere enten fysisk kuttet cellen ut av sitt omkringliggende vev gjennom laserfangstmikrodisseksjon (LCM)27 eller utført enzymatisk fordøyelse av celleveggene28. Blant de sistnevnte cellene, såkalte protoplasts, er befolkningen av interesse fluorescerende merket og kan skilles via fluorescensaktivert cellesortering (FACS)7. LCM krever vanligvis en prøve som skal festes og bygges inn i voks, noe som til slutt forverrer kvaliteten på RNA29. FACS-baserte metoder gir høy kvalitet RNA, men prosessen med å protopelastere i seg selv introduserer forskjeller i genuttrykk30 og vev med modifiserte og tykke sekundære cellevegger er notorisk vanskelig å behandle. Videre antas mange utviklingsprosesser i planter å stole på mekanisk overførte signaler, og derfor er integriteten til celleveggen av avgjørende betydning31. To metoder, som bruker en snarvei til å omgå celleisolasjon ved å operere på nivået av kjerner, er fluorescensaktivert kjernefysisk sortering (FANS) og isolering av kjerner merket i bestemte celletyper (INTAKT). Som i TRAP bruker de celletypespesifikke arrangører for å markere kjerner, som senere blir beriket via sortering eller pull down, henholdsvis8,15. En stor utfordring for alle disse tilnærmingene er å få tilstrekkelig RNA-materiale fra undergrupper av celler i et vev. Som TRAP fanger bare en brøkdel av cellulære RNAs, prøvesamling er en betydelig flaskehals. Spesielt sensitive bibliotekforberedelsesprotokoller er derfor nødvendig for å produsere data av høy kvalitet fra lave inngangsbeløp.
Siden etableringen har TRAP enten blitt brukt i kombinasjon med DNA-mikroarrayer eller, som sekvenseringskostnader falt betydelig de siste årene, RNA-seq10,32,33. En rekke forskningsspørsmål har allerede blitt belyst som gjennomgått i Sablok et al.34. Vi er overbevist om at flere rapporter vil følge i de kommende årene som teknikken er svært allsidig når du kombinerer ulike arrangører for å målrette bestemte celletyper. Til slutt vil dette bli gjort selv på en utilbørlig måte, og kan kombineres med å undersøke anleggets reaksjon på mange biotiske og abiotiske stressfaktorer. I tillegg, der stabile transgene linjer ikke er tilgjengelige, har hårete rotuttrykkssystemer også blitt brukt til å utføre TRAP i tomat og medicago35,36.
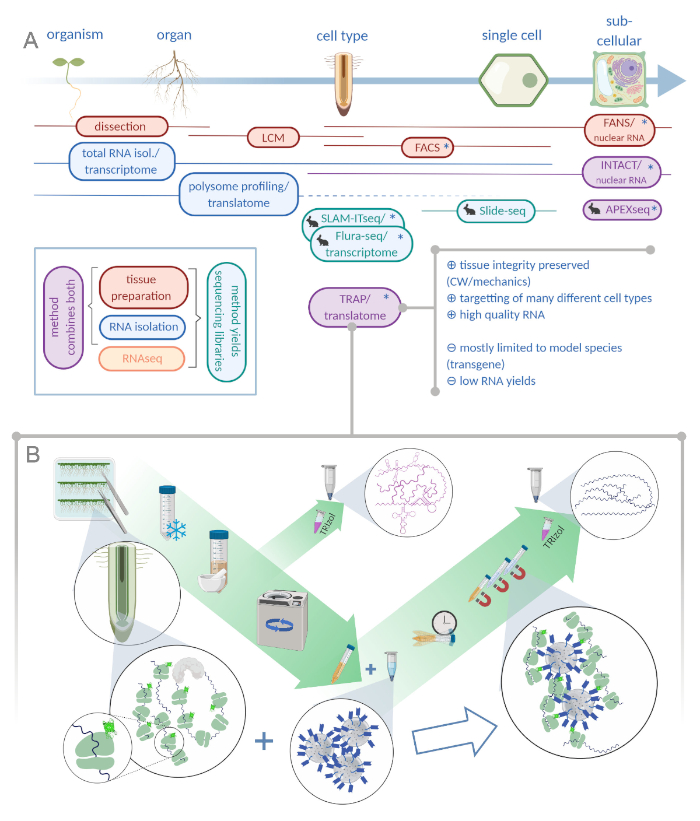
Figur 1: Translating ribosome affinity purification (TRAP) utfyller “omics” analyseporteføljen. A. Økende nivåer av analytisk presisjon, ned til encellede eller til og med subcellulær oppløsning kan oppnås ved en mengde metoder eller kombinasjoner av disse. Ordningen gir en oversikt over tilgjengelige verktøy i plante- og dyrefeltet. Vevsinnsamling ved cellulær oppløsning kan oppnås ved protokoller som LCM eller FACS, som deretter kobles til standard transkripsjon eller polysome profilering / translatome analyse. TRAP og INTAKT integrerer både vevsfangst og RNA-isolasjon som de er basert på epitop-tagging. Intakte prøver bare cellekjerner og utgjør derfor et spesielt tilfelle av transkripsjonsanalyse. Et lite kaninikon markerer nyutviklede metoder i dyrefeltet: Mens SLAM-ITseq og Flura-seq er avhengige av metabolsk targetting av nascent RNAs med modifiserte uracilbaser i celler som uttrykker det tillatte enzymet, bruker Slide-seq et belagt glasslysbilde med DNA-strekkoder som gir posisjonsinformasjon i cellulærområdet. En nærhetsmerkingstilnærming følges i APEX-seq for å prøve RNAer i bestemte undercellulære rom. Spesielt krever økt oppløsning ofte generering av transgen materiale (stjerner) og disse metodene brukes dermed hovedsakelig til modellarter. TRAP er spesielt egnet for plantevitenskapsstudier som involverer cellevegg (CW) eller mekanikersignalering samt cellearter som er vanskelige å frigjøre fra cw-matrisen. B. Detaljerte våtlabtrinn i TRAP-prosedyren: Frøplanter som uttrykker GFP-merket ribosomalt protein i forskjellige celletyper (f.eks. rotendodermis) dyrkes på Petri-retter i syv dager og rotmateriale høstet ved snap frysing. En total RNA-kontrollprøve samles inn fra det homogeniserte råoljeekstraktet før ruskene blir avstøtende via sentrifugering. Magnetiske anti-GFP perler legges til det klarerte ekstraktet for å utføre immunnedbør. Etter inkubasjon og tre vasketrinn oppnås polysomassosiert RNA (TRAP/polysome RNA) direkte via fenol-kloroformekstraksjon. LCM: laser fangst mikrodisseksjon, FACS / FANS: fluorescens-aktivert celle / kjernefysisk sortering, APEX-seq: metode basert på konstruert ascorbat peroksidans, INTAKT: isolering av kjerner merket i spesifikke celletyper, SLAM-ITseq: tiol (SH)-koblet alkylation for metabolsk sekvensering av RNA i vev, Flura-seq: fluorouracil-merket RNA sekvensering (Opprettet med Biorender.com) Vennligst klikk her for å se en større versjon av denne figuren.
Målet med denne artikkelen er å gi en detaljert beskrivelse av TRAP-metoden, for å markere kritiske trinn og gi veiledning for en mulig bibliotekklargjøringsmetode.
Et generisk TRAP-eksperiment vil i hovedsak bestå av følgende trinn (se også figur 1B): (1) Tilberedning av plantemateriale, inkludert kloning av ribosom-tagging konstruksjon, transgen linjeproduksjon og valg, voksende og bulking opp av frø, sterilisering og plating, og stress søknad / behandling (valgfritt) og vev høsting; (2) immunrensing inkludert vevhomogenisering og rydding av råekstrakt, perlevask og immunrensing, og vasketrinn; (3) RNA-ekstraksjon og kvalitetsvurdering; og (4) bibliotekforberedelse.
Arabidopsis roten har vært et modellsystem for å studere planteutvikling helt siden introduksjonen som modellanlegg37,38. Her vises anvendelsen av TRAP i sammenheng med plantesiderotutvikling. I planter er oppbyggingen av hele rotsystemet avhengig av utførelsen av dette programmet og er derfor svært viktig for organismens overlevelse39. I Arabidopsisstammer laterale røtter fra pericycle vev som ligger ved siden av xylem fartøy og derfor kalles xylem pole pericycle (XPP; se figur 2C)40. Noen XPP-celler, som ligger dypt inne i roten, får en grunnleggercelleidentitet og, på en lokal hormonell utløser, begynner å spre seg ved hevelse og dele anticlinally41. Men på grunn av tilstedeværelsen av en stiv celleveggmatrise, utøver denne prosessen mekanisk stress på det omkringliggende vevet. Spesielt påvirkes den overliggende endodermis, da den er i veien for den laterale rotvekstaksen42,43,44. Faktisk må det nylig dannede primordium vokse gjennom den overliggende endodermiscellen (Figur 2C2) mens cortex- og epidermis-celler bare skyves til side for primordium å endelig dukke opp45,46. Nylig arbeid i laboratoriet vårt har vist at endodermis aktivt bidrar til å imøtekomme spredningen i perisyklusen. Målrettet blokkering av endodermal hormonell signalisering er tilstrekkelig til å hemme selv den aller første divisjonen i XPP-cellene47. Derfor utgjør pericycle-endodermis kommunikasjon et svært tidlig kontrollpunkt for lateral rotutvikling i arabidopsis. Det er imidlertid ikke kjent hvordan denne crosstalk utføres. For å løse dette mysteriet valgte vi TRAP-seq-tilnærmingen for å målrette XPP og endodermale celler. For å berike for celler i siderotprogrammet, etterlignet vi den hormonelle utløseren ved å eksogene bruke en auxin analog (1-naftaleneddiksyre, NAA)48, som samtidig lov til å midlertidig løse den første fasen av lateral rotdannelse.
