Driven av den ökande tillämpningen av nästa generations sekvenseringstekniker skulle rumslig upplösning i utvecklingsbiologi kunna utökas. Samtida studier syftar till att dissekera vävnader ner till specialiserade celltyper, om inte encellig nivå1,,2,,3,4. I detta syfte har en uppsjö av olika metoder utarbetats under de senaste femtio åren (se figur 1A)5,,6,,7,,8,,9,,10,,11,,12,,13,,14,,15.
Många verktyg inom växtvetenskap har varit anpassningar av tekniker som var pionjärer inom djurforskning. Detta är inte fallet för den metod som vi inför i detalj här. Under 2005, utrustad med en stark bakgrund inom proteinöversättning, Bailey-Serres Lab syftar till att ingenjör ribosomala proteiner för efterföljande affinitet rening16. Således kunde de undvika tidskrävande och arbetsintensiv polysome profilering, som bygger på ultracentrifugering med en sackaros gradient och användes för att bedöma översätta ribosomer sedan 1960-talet17,18. Metoden har sedan dess kallat translationell ribosomaffinitetsrening (TRAP)16. Efter framgångsrika översättarstudier på växter, Heiman et al. anpassade TRAP för djur19 och andra utvidgas dess tillämpning till jäst20, Drosophila21, Xenopus22 och zebrafisk23,24.
Även om genetisk modifiering av modellsystemet är en förutsättning för TRAP, som begränsar dess tillämpning till arter som kan användas för genetisk omvandling, kan man samtidigt utnyttja denna invändning mot målundergrupper av celler som är av särskilt intresse och i övrigt extremt svårt att isolera från intakt vävnad/organ25 (t.ex. höggrenade dendritiska celler i en mushjärna eller svamphyfa i infekterad växtvävnad). I växter hålls alla celler på plats via cellväggar som ligger till grund för det hydrostatiska skelettet26. För att frigöra en växtcell från denna matris, forskare har antingen fysiskt skära cellen ur sin omgivande vävnad genom laser fånga microdissection (LCM)27 eller utförs enzymatisk matsmältning av cellväggarna28. Bland de senare cellerna, så kallade protoplaster, är populationen av intresse fluorescerande märkt och kan separeras via fluorescensaktiverad cellsortering (FACS)7. LCM kräver vanligtvis ett prov som skall fastställas och inbäddad i vax, vilket i slutändan försämrar kvaliteten på dess RNA29. FACS-baserade metoder ger högkvalitativa RNA, men processen att protoplasting själv introducerar skillnader i genuttryck30 och vävnader med modifierade och tjocka sekundära cellväggar är notoriskt svåra att behandla. Dessutom antas många utvecklingsprocesser i anläggningar förlita sig på mekaniskt överförda signaler och därför är cellväggens integritet av största vikt31. Två metoder, som använder en genväg för att kringgå cellisolering genom att arbeta på nivån av kärnor, är fluorescensaktiverad kärnsortering (FANS) och isolering av kärnor taggade i specifika celltyper (INTAKT). Liksom i TRAP använder de celltypspecifika promotorer för att markera kärnor, som därefter berikas via sortering eller pull down,respektive 8,15. En stor utmaning för alla dessa metoder är att få tillräckligt RNA-material från delmängder av celler i en vävnad. Som TRAP fångar bara en bråkdel av den cellulära RNAs, prov insamling är en betydande flaskhals. Därför behövs särskilt känsliga biblioteksförberedelseprotokoll för att producera högkvalitativa data från låga indatamängder.
Sedan starten har TRAP antingen använts i kombination med DNA-mikroarrayer eller, som sekvensering kostnader minskat betydligt under de senaste åren, RNA-seq10,32,33. En mängd forskningsfrågor har redan klarlagts som granskats i Sablok et al.34. Vi är övertygade om att fler rapporter kommer att följa under de kommande åren eftersom tekniken är mycket mångsidig när man kombinerar olika initiativtagare för att rikta specifika celltyper. Så småningom, Detta kommer att ske även på ett inducerande sätt, och kan kombineras med sondering av anläggningens reaktion på många biotiska och abiotiska stressfaktorer. Dessutom, där stabila transgena linjer inte finns tillgängliga, håriga rot uttryckssystem har också framgångsrikt använts för att utföra TRAP i tomat och medicago35,36.
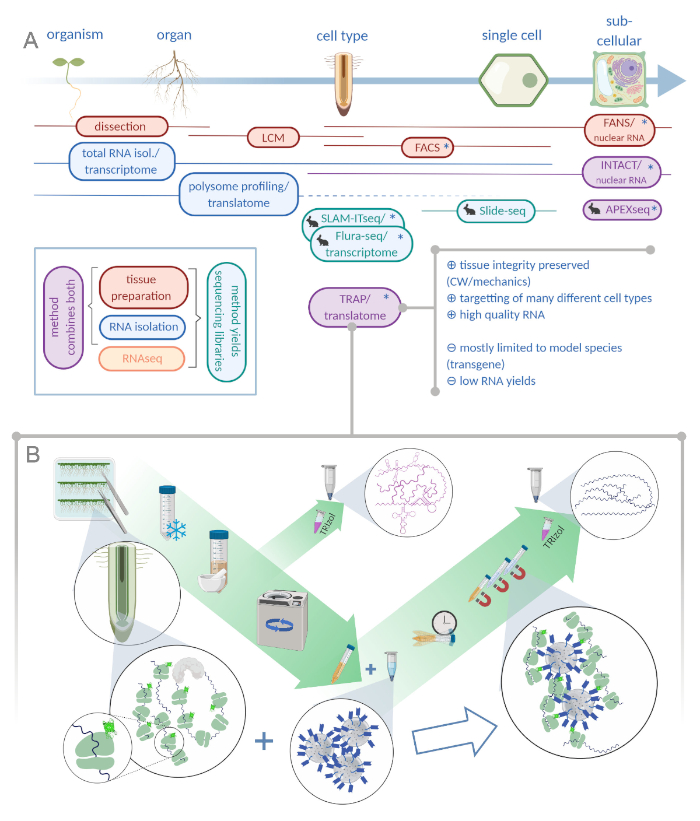
Figur 1: Omräkning av ribosomaffinitetsrening (TRAP) kompletterar analysportföljen “omics”. A. Ökande nivåer av analytisk precision, ner till encellig eller till och med subcellulär upplösning kan uppnås genom en uppsjö av metoder eller kombinationer av dessa. Systemet ger en översikt över de verktyg som för närvarande finns tillgängliga på växt- och djurområdet. Vävnadssamling vid cellupplösning kan uppnås genom protokoll som LCM eller FACS, som sedan kopplas till standard transkriptom eller polysome profilering / översättaranalys. TRAP och INTACT integrerar både vävnadsfångst och RNA-isolering eftersom de är baserade på epitop-märkning. Intakt prover endast cellkärnor och utgör därför ett specialfall av transkriptom analys. En liten kanin ikon markerar nyutvecklade metoder inom djurområdet: Medan SLAM-ITseq och Flura-seq förlita sig på metabolisk targetting av begynnande RNAs med modifierade uracil baser i celler som uttrycker tillåtande enzym, Slide-seq använder sig av en bestruken glas bild med DNA-streckkoder som ger positionsinformation i cellområdet. En närhetsmärkning följs i APEX-seq för att prova RNA i specifika subcellulära fack. Framför allt kräver ökad upplösning ofta generering av transgent material (asterisker) och dessa metoder används därför främst för modellarter. TRAP är särskilt lämpad för växtvetenskapliga studier med cellvägg (CW) eller mekaniker signalering samt cellarter som är svåra att släppa från sin CW matris. B. Detaljerade våt-lab steg i TRAP förfarandet: Plantor som uttrycker GFP-märkt ribosomalt protein i olika celltyper (t.ex. rot endodermis) odlas på petriskålar i sju dagar och rotmaterial skördas genom snap frysning. Ett totalt RNA-kontrollprov samlas in från det homogeniserade råextraktet innan skräpet pelleteras via centrifugering. Magnetiska anti-GFP pärlor läggs till rensade extrakt för att utföra immunprecipitation. Efter inkubation och tre tvättsteg erhålls det polysome-associerade RNA (TRAP/polysome RNA) direkt via fenol-kloroformextraktion. LCM: mikroavskiljning av laseravskiljning, FACS/FLÄKTAR: fluorescensaktiverad cell-/kärnsortering. APEX-seq: metod baserad på konstruerad askorbatperoxidas, INTAKT: isolering av kärnor taggade i specifika celltyper, SLAM-ITseq: thiol(SH)-linked alkylation för metabolisk sekvensering av RNA i vävnad, Flura-seq: fluorouracil-märkt RNA-sekvensering (Skapad med Biorender.com) Klicka här för att se en större version av denna figur.
Målet med den här artikeln är att tillhandahålla en detaljerad beskrivning av TRAP-metoden, att markera kritiska steg och att ge vägledning för en möjlig biblioteksförberedelsemetod.
Ett generiskt TRAP-experiment kommer huvudsakligen att bestå av följande steg (se även figur 1B):(1) Beredning av växtmaterial, inklusive kloning av ribosommärkningskonstruktion, transgen linjeproduktion och val, odling och sammanslagning av frön, sterilisering och plätering samt stressapplicering/behandling (frivillig uppgift) och vävnadsskörd. (2) immunrening inklusive vävnad homogenisering och clearing av rå extrakt, pärla tvätt och immunrening, och tvätta steg; (3) RNA-utvinning och kvalitetsbedömning. och (4) biblioteksförberedelser.
Den Arabidopsis roten har varit en modell system för att studera växtutveckling ända sedan den infördes som en modell anläggning37,38. Här visas tillämpningen av TRAP upp i samband med växt lateral rot utveckling. I växter, uppbyggnaden av hela rotsystemet bygger på genomförandet av detta program och är därför mycket viktigt för överlevnaden av organismen39. I Arabidopsiskommer laterala rötter från pericycle vävnad som finns bredvid xylem fartyg och därför kallas xylem pole pericycle (XPP; se figur 2C)40. Vissa XPP celler, som ligger djupt inne i roten, förvärva en grundare cell identitet och, på en lokal hormonell trigger, börjar föröka sig genom svullnad och dela anticlinally41. Men på grund av närvaron av en styv cellvägg matris, utövar denna process mekanisk stress på de omgivande vävnaderna. I synnerhet påverkas den överliggande endodermis, som det är i vägen för den laterala rottillväxtaxeln42,43,44. Faktum är att den nybildande primordium måste växa genom den överliggande endodermis cellen (Figur 2C2) medan cortex och epidermis celler bara skjuts åt sidan för primordium att äntligen dyka upp45,46. Senaste arbetet i vårt labb har visat att endodermis aktivt bidrar till att tillgodose spridningen i pericycle. Riktad blockering av endodermal hormonell signalering är tillräcklig för att hämma även den allra första divisionen i XPP-cellerna47. Därför utgör pericycle-endodermis kommunikation en mycket tidig kontrollpunkt för lateral rot utveckling i Arabidopsis. Det är dock inte känt hur denna överhörning utförs. För att reda ut detta mysterium valde vi TRAP-seq-metoden för att rikta in sig på XPP och endodermala celler. För att berika för celler i det laterala rotprogrammet, härmade vi den hormonella utlösaren genom att exogent tillämpa en auxin analog (1-naftalenetiksyra, NAA)48, som samtidigt tillåtet att tidsmässigt lösa den inledande fasen av laterala rotbildning.
