Reactive oxygen species (ROS) signalering reguleert de ontwikkeling en werking van het zenuwstelsel1. Een belangrijke cellulaire ROS-bron zijn NADPH-oxidasen (NOX), transmembrane-eiwitten die superoxide en waterstofperoxide genereren (H2O2)2. NOX-enzymen worden gevonden in het centrale zenuwstelsel (CZS) en NOX-afgeleide ROS dragen bij aan neuronale ontwikkeling3,4,5,6. Onderhoud en differentiatie van neurale stamcellen, het vaststellen van neuronale polariteit, neurietuitgroei en synaptische plasticiteit hebben voldoende ros7,8,9,10,11nodig . Aan de andere kant draagt ongecontroleerde productie van ROS door NOXen bij aan neurodegeneratieve aandoeningen, waaronder de ziekte van Alzheimer, multiple sclerose en traumatisch hersenletsel12,13,14. Daarom is de productie van fysiologisch relevante ROS van cruciaal belang voor het behoud van gezonde omstandigheden.
De ontwikkeling van genetisch gecodeerde biosensoren vergemakkelijkte de detectie van cellulaire ROS aanzienlijk. Een belangrijk voordeel van genetisch gecodeerde biosensoren is de verhoogde temporele en ruimtelijke resolutie van het ROS-signaal, omdat deze sensoren specifiek op verschillende locaties kunnen worden gericht. Redox-gevoelige GFP (roGFP) is een type van dergelijke ROS-biosensoren. De roGFP2-Orp1-variant detecteert specifiek H2O2 via zijn Orp1-domein, een glutathion peroxiredoxinefamilie-eiwit uit gist15,16. De oxidatie van het Orp1-eiwit wordt overgebracht naar roGFP2 om de conformatie ervan te wijzigen (figuur 1A). De sonde vertoont twee excitatiepieken in de buurt van 405 nm en 480 nm en een enkele emissiepiek bij 515 nm. Bij oxidatie verandert de fluorescentie-intensiteit rond excitatiepieken: terwijl 405 nm excitatie toeneemt, neemt 480 nm excitatie af. RoGFP2-Orp1 is dus een ratiometrische biosensor en H2O2-niveaus worden gedetecteerd door de verhouding van fluorescentieintensiteiten opgewekt op twee verschillende golflengten (Figuur 1B). Over het algemeen is roGFP2-Orp1 een veelzijdig hulpmiddel voor ROS-beeldvorming dat efficiënt in vivokan worden gebruikt.
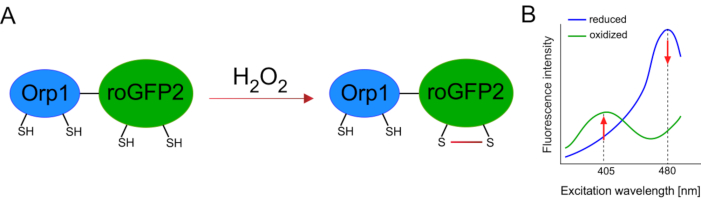
Figuur 1: Schematische representatie en excitatiespectra van roGFP2-Orp1. (A) Oxidant transfer vindt plaats tussen Orp1 en roGFP2 als reactie op H2O2, wat leidt tot conformationele veranderingen in roGFP2. (B) De excitatiespectra van de roGFP2-Orp1 vertoont twee excitatiepieken bij 405 nm en 480 nm en enkele emissiepiek bij 515 nm. Bij oxidatie door H2O2neemt de excitatie van 405 nm toe terwijl de excitatie van 480 nm afneemt. Dit resulteert in een ratiometrische uitlezing voor H2O2 aanwezigheid. Het cijfer is gewijzigd van Bilan en Belousov (2017)16 en Morgan et al. (2011)25. Klik hier om een grotere versie van deze afbeelding te bekijken.
Het Danio rerio (zebravis) modelsysteem heeft verschillende voordelen voor het toepassen van genetisch gecodeerde biosensoren. De optische transparantie van de embryo’s en larven maakt niet-invasieve in vivo beeldvorming mogelijk. Er worden nieuwe beeldvormingstools ontwikkeld om een hogere resolutie en diepere penetratie te bereiken17. Verder zijn er gevestigde instrumenten voor genetische manipulatie (ectopische mRNA-expressie, Tol2-transgenese, enz.) en genoombewerking (TALEN’s, CRISPR/Cas9, enz.), die de generatie van transgene dierenbevordert 18. Naarmate de zebravisembryo’s zich buiten de moeder ontwikkelen, maakt dit systeem verder gemakkelijkere toegang tot en manipulatie van de embryo’s mogelijk. Bijvoorbeeld, eencellige stadium injecties en medicamenteuze behandelingen kunnen gemakkelijk worden gedaan.
Hier gebruikten we zebravissen om de H 2O2-specifiekebiosensor roGFP2-Orp1 tijdelijk uit te drukken door in vitro getranscribeerd mRNA te injecteren. Deze embryo’s kunnen worden gebruikt voor zowel in vitro beeldvorming van gekweekte neuronen als in vivo beeldvorming (figuur 2). We beschrijven een protocol voor het ontleden en plateren van retinale ganglioncellen (RGCs) uit zebravisembryo’s, gevolgd door het beoordelen van H2O2-niveaus in gekweekte neuronen. Vervolgens presenteren we een methode voor in vivo beeldvorming van roGFP2-Orp1-uitdrukkende embryo’s en larven met behulp van confocale microscopie. Deze aanpak maakt het niet alleen mogelijk om fysiologische H2O2-niveauste bepalen, maar ook potentiële veranderingen die optreden in verschillende ontwikkelingsstadia of omstandigheden. Over het algemeen biedt dit systeem een betrouwbare methode voor het detecteren van H2O2 in levende cellen en dieren om de rol van H2O2 in ontwikkeling, gezondheid en ziekte te bestuderen.
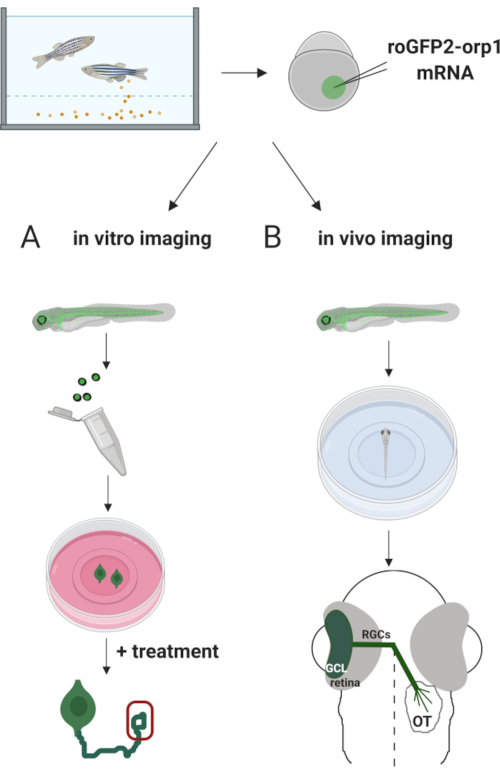
Figuur 2. Schets van de experimentele aanpak. Kort na het verzamelen van embryo’s wordt roGFP2-Orp1 mRNA geïnjecteerd in de dooier van zebravisembryo’s in één celstadium. Het ontwikkelen van embryo’s kan worden gebruikt voor zowel (A) in vitro als (B) in vivo beeldvorming. (A) GFP-positieve embryo’s worden gebruikt om netvlies te ontleden voor RGC-verzameling bij 34 hpf. Gedissocieerde RGCs worden geplateerd op PDL/laminine-gecoate coverslips in ZFCM (+) media. Groeikegelbeeldvorming kan worden uitgevoerd terwijl RGC’s hun axonen verlengen na 6-24 uur plateren. Cellen kunnen worden onderworpen aan verschillende behandelingen om de potentiële veranderingen in H 2 O2-niveauste meten. Hier hebben we H 2 O2-niveausgemeten in de groeikegels van RGCs (rood). (B) GFP-positieve embryo’s worden gebruikt voor in vivo beeldvorming. Op de gewenste leeftijd kunnen embryo’s worden verdoofd en gemonteerd op glazen bodemschalen van 35 mm voor confocale beeldvorming. Hier worden embryo’s ventrally gemonteerd voor retinale beeldvorming. Schematisch toont retinale ontwikkeling bij zebravissen. RGCs vormen ganglioncellaag (GCL), de binnenste laag in het netvlies. RGC axonen ontwikkelen zich tot oogzenuw om de middellijn te kruisen en vormen optisch chiasme. Vervolgens groeien RGC axonen dorsaal om synapsen te maken in het optische tectum in de middenhersenen. Klik hier om een grotere versie van deze afbeelding te bekijken.
