Primary culture of astrocytes infected with LV-G1-MitoTimer exhibited typical mitochondrial networks. Before treatment, astrocytes expressing LV-G1-MitoTimer showed the heterogeneous mitochondrial size and various green/red fluorescence intensities (Figure 3, Figure 4, and Video 1). The mitochondrial system of astrocyte cultures expressing LV-G1-MitoTimer was monitored before and after incubation with H202 (10 µM). The different mitochondrial features described above were calculated over 12 h (every 3 h) and normalized (cell by cell) to their initial state. At the morphological level (Figure 3B), the effects of H2O2 start to be visible at about 6 h. Indeed, the mitochondria were fragmented (decrease of length, surface area, and elongation factor). This fragmentation is even more obvious 12 h after the treatment. Note that the diameters, widths, and sphericity were not reduced. Concerning redox state and turnover (Figure 3C), 3 h after H2O2 treatment, the proportion of green mitochondria increased in astrocytes (the consequence of a rapid increase in mitochondrial biogenesis). At 6 h, the green/red ratio returned to baseline levels, but the number of purely red mitochondria increased significantly from basal levels. After 12 h, the consequences of the oxidative treatment of H2O2 were visible and resulted in a substantial increase in the ratio and number of red puncta. Concerning the dynamics and mobility (Figure 3D), 3 h after the treatment, all the criteria were transiently increased. In the longer term (12 h), the mitochondria moved more slowly and over shorter distances.

Figure 1: Astrocyte culture expressing LV-G1-MitoTimer biosensor. (A) Confocal photographs of astrocytes expressing LV-G1-MitoTimer. (B) Selection of confocal photographs of reduced (green) balanced (orange) and oxidized (red) mitochondria with different levels of fragmentation. (C) Summary diagram of the different criteria available for analysis in an astrocyte expressing LV-G1-MitoTimer. Scale bar: (A) Upper panel: 50 µm, lower panel: 10 µm, (B) 1 µm. Please click here to view a larger version of this figure.
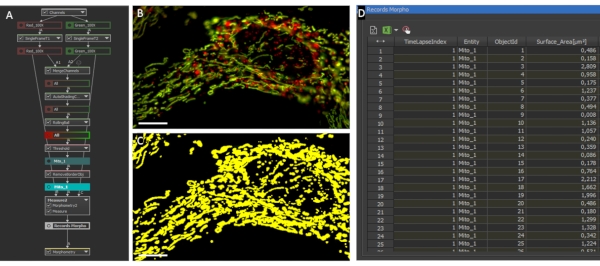
Figure 2: Mitochondrial morphology and ratio analysis. (A) GA3 script overview for the analysis of individual mitochondrial morphology and ratio. (B) Initial photographs of an astrocyte expressing LV-G1-MitoTimer analyzed with the GA3 script. (C) Example of binary masks generated for the mitochondrial system of astrocytes. Scale bar: 10 µm (B-C). Please click here to view a larger version of this figure.
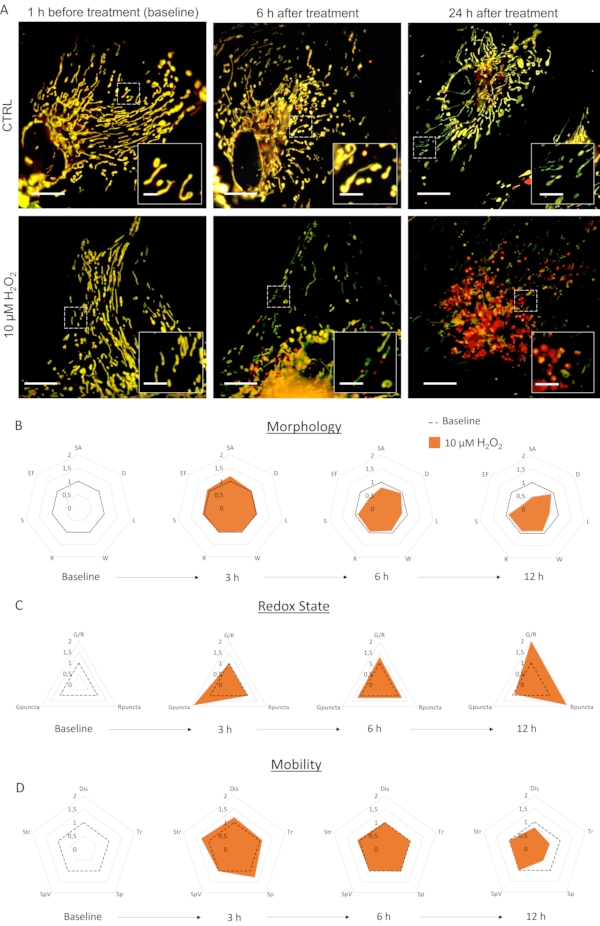
Figure 3: The effects of H2O2 on the mitochondrial system of astrocytes. (A) Photographs of astrocytes expressing LV-G1-MitoTimer 1 h before and 6 h, 24 h after treatment with PBS (CTRL) and 10 µM of H2O2. (B) Radar charts of mitochondrial morphology, (C) redox state and turnover, and (D) mobility criteria evaluated on astrocytes during baseline and 3 h, 6 h, and 12 h after H2O2 treatment. SA: Surface area; D: Diameter; L: Length; W: Width; R: Roundness; S: Sphericity; EF: Elongation factor (=L/W); G/R: Individual red/green ratio; Gpuncta: Percentage of green puncta mitochondria; Rpuncta: Percentage of red puncta mitochondria; Dis: Displacement; Tr: Track Length; Sp and SpV: Speed and speed variance; Str: Straightness. Scale bar: 20 µm (A) and 2.5 µm (inset). Please click here to view a larger version of this figure.
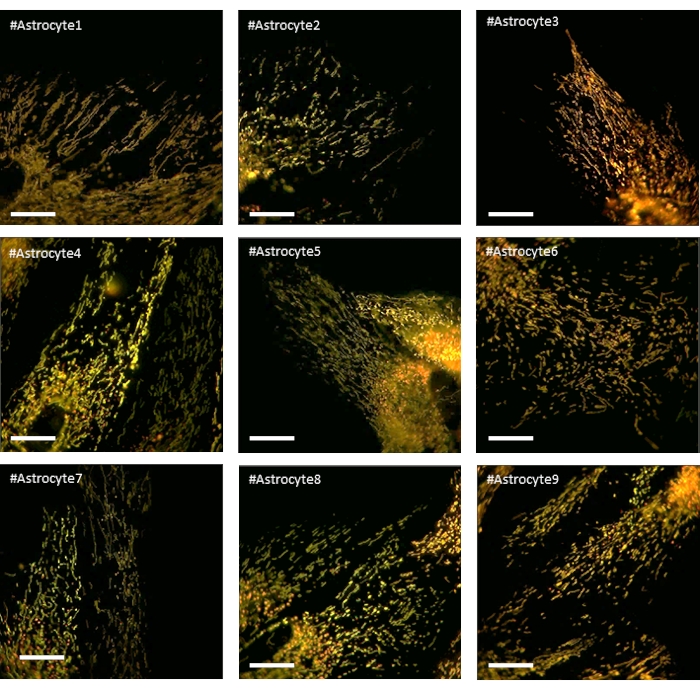
Figure 4: Photographs of astrocytes expressing LV-G1-MitoTimer and showing a homogeneous and balanced mitochondrial network during baseline. Scale bar: 20 µm Please click here to view a larger version of this figure.
Video 1: Effect of H2O2 treatment on the mitochondrial system of cultured astrocytes. Astrocytic mitochondria before H2O2 treatment (baseline), as well as 6 h and 24 h after H2O2 treatment compared to non-treated control cell. Please click here to download this File.
| Morphology criteria | Range | Remarks | ||
| Surface Area (SA) | 0.5–4 µm2 | These criteria inform on the fragmented-elongated features of mitochondria. They generally evolve in the same direction. Fragmented mitochondria will have decreased surface area, diameter, length, and elongation factor while roundness, sphericity, and width may be unchanged or increased. | ||
| Diameter (D) | 0.5–1.5 µm | |||
| Length (L) | 0.5–5 µm | |||
| Width (W) | 0.5–2 µm | |||
| Roundness (R) | 0–1 | |||
| Sphericity (S) | 0–1 | |||
| Elongation factor (EF = L/W) | 1–10 | |||
Table 1: Summary of selected parameters for mitochondrial morphology.
| Redox State criteria | Range | Remarks | ||
| Individual ratio (G/R) | 0–10 | The ratio indicates the result of the redox state. It informs about the general state and age of the mitochondria in the cell. It is essential to consider that this ratio is the balance of biogenesis and degradation of mitochondria and the fission/fusion of oxidized mitochondria with reduced mitochondria. Therefore the evaluation of the number of green and red puncta can powerfully help interpret the results. Green puncta mitochondria are determined when the intensity of green is 10 times that of red. Red puncta mitochondria are determined when the intensity of red is 10 times greater than that of green. The redox state of an astrocyte is the average of all the mitochondria ratios of that cell. | ||
| Percentage of green puncta mitochondria (Gpuncta) | 0%–100% | |||
| Percentage of red puncta mitochondria (Rpuncta) | 0%–100% | |||
Table 2: Summary of selected parameters for mitochondrial redox state.
| Mobility criteria | Range | Remarks | ||
| Displacement (Dis) | 0–10 µm | Together these features inform the general motility dynamics of the network. Stationary mitochondria display short displacement & track length with a low speed. On the other hand, oscillatory particles can be differentiated with a difference between the track length and displacement (resulting in low straightness) and an increased speed compared to static. | ||
| Track Length (Tr) | 0–10 µm | |||
| Speed and speed variance (Sp and SpV) | 0–1.5 µm/s ± 0.2 µm/s | |||
| Straightness | 0–1 | |||
| (Str = displacement/track length) | ||||
Table 3: Summary of selected parameters for mitochondrial mobility.
Supplemental Coding File 1: GA3 script file for analysis of individual mitochondrial morphology. Please click here to download this File.
