מודל העכברים של ניוון עצבי שנותח כאן עשוי לסייע בחקר היבטים קריטיים רבים המעורבים בפתופיזיולוגיה של מחלת פרקינסון, כולל המנגנונים המעורבים בפתולוגיה של αSyn והפעלה מיקרוגליאלית, המעורבות של מערכת החיסון ההיקפית בוויסות של דלקת עצבית, ומנגנוני הניוון העצבי. בין המנגנונים המעורבים בפתולוגיה של αSyn הם אותם מנגנונים תת-תאיים הקשורים לתפקוד מיטוכונדריאלי, ליזוזומלי או פרוטאסומלי בנוכחות עומס מוגזם של αSyn בתאי העצב הדופמינרגיים של SN2. חשוב לקחת בחשבון שבנוסף לביטוי hαSyn המושרה על ידי התמרה בתיווך AAV, העכבר האנדוגני αSyn תורם גם הוא לעומס של ביטוי αSyn הכולל. עכברים מהונדסים המבטאים יתר על המידה עכבר αSyn מפתחים פתולוגיה סינוקלאינית דומה, נוירופתולוגיה ופגיעה מוטורית למודלים של עכברים המבוססים על ביטוי יתר של hαSyn32. בנוגע להפעלה מיקרוגליאלית, מודל העכבר הנוכחי עשוי לשמש כדי לחקור כיצד שחקנים מולקולריים ותאיים שונים כגון ציטוקינים, נוירוטרנסמיטורים, אסטרוציטים, נוירונים, מחסום הדם-מוח ותאי T עשויים לווסת את הרכישה של פנוטיפים פונקציונליים פרו-דלקתיים או אנטי-דלקתיים 8,10,11 . מודל זה מהווה גם כלי חשוב לחקר תפקידה של מערכת החיסון ההיקפית, כולל לא רק תאי T אלא גם מקרופאגים, מונוציטים ונייטרופילים, על תהליכי הדלקת העצבים והניוון העצבי של נוירונים ניגרליים 11,33,34. לבסוף, מודל עכבר זה מייצג גם מערכת רבת ערך לחקר המנגנונים התאיים והמולקולריים של ניוון עצבי in vivo, כולל אלה המושרים על ידי תהליכים תאיים פנימיים, כגון עקה חמצונית, ליקויים באנרגיה ואברונים פגומים 2, או אלה המופעלים על ידי שחקנים חיצוניים, כגון גורמים נוירוטוקסיים המיוצרים על ידי תאים מיקרוגליאליים, אסטרוציטים ותאי T ציטוטוקסיים8, 28,29,35.
מגבלה של מודל עכבר זה היא המחקר של האופן שבו הצבירה הפתולוגית של αSyn במיקומים חוץ-מוחיים עשויה להוות את השלבים הראשונים בהתפתחות מחלת פרקינסון36. בהקשר זה, ישנן עדויות הולכות וגוברות המצביעות על כך שלפני הניוון העצבי של נוירונים ניגרליים ופגיעה מוטורית, הפתולוגיה של αSyn מתחילה ברירית המעיים ובאפיתל חוש הריח36, וככל הנראה גם בתגובת תאי ה-T הספציפיים ל-αSyn12. לאחר מכן, אגרגטים של αSyn היו נודדים דרך עצב הוואגוס אל גזע המוח, ומפעילים את הדלקת העצבית והניוון העצבי של נוירונים דופמינרגיים12. למרות שמודל AAV-hαSyn משחזר את רוב ההיבטים של מחלת פרקינסון, אין מעורבות ניכרת של הצבירה הפתולוגית של αSyn במיקומים חוץ-מוחיים במודל זה. מודל חלופי הכולל פתולוגיה של hαSyn המתאים לחקר היבטים אלה של מחלת פרקינסון עשוי להיות עכברים מהונדסים המבטאים יתר על המידה את hαSyn תחת שליטתו של מקדם Thy1, מודל Thy1-SNCA37, שבו התפתחות המחלה תלויה במיקרוביוטה של המעיים וכוללת פגיעה ניכרת במערכת העיכול38.
למרות שזה מועיל לחקר התהליכים המגוונים הקשורים לפתופיזיולוגיה של מחלת פרקינסון, מודל העכבר הנוכחי כולל צעדים קריטיים שיש לבדוק בדקדקנות, כולל העברה נכונה של הווקטורים הנגיפיים בקואורדינטות המרחביות המתאימות, הביטוי הסלקטיבי של hαSyn בתאי עצב (התלוי בסרוטיפ AAV ובמבנה הווקטורי), ומינון ותזמון AAV תקינים לפני ניתוח הפנוטיפ הפרקינסון. ניתוח המסירה הנכונה של הווקטורים הנגיפיים ב- SN הוא הכרחי, שכן השימוש בקואורדינטות המרחביות הנכונות של ה- SN עשוי שלא להספיק כאשר המחט אינה ישרה לחלוטין, שלעתים אינה מורגשת לעין האנושית. יתר על כן, הדיפוזיה של וקטורי AAV תלויה בסרוטיפ AAV39. מסיבות אלה, יש צורך לבצע בקרות איכות תקופתיות הבודקות את המסירה הנכונה והדיפוזיה של וקטורי AAV-GFP המוזרקים בעקבות תצפית של GFP בפרוסות מוח המכילות את שטח ה- SN.
לגבי הביטוי הסלקטיבי של hαSyn בתאי עצב, באופן עקרוני, הביטוי של hαSyn יכול להיות מהונדס להיות נשלט על ידי מקדם סלקטיבי עבור נוירונים או, אפילו יותר מדויק, סלקטיבי עבור נוירונים דופמינרגיים, כגון השימוש של מקדם TH בווקטורים AAV כדי לגרום לביטוי סלקטיבי של גנים בתאי עצב דופמינרגיים40 . עם זאת, אסטרטגיה זו אינה פועלת כאשר מה שמתבקש הוא ביטוי יתר של הגן של העניין. מסיבה זו, במודל הנוכחי, חיוני להשתמש במקדם חזק (מקדם הגורם לביטוי גבוה של הגן במורד הזרם) ובסרוטיפים של AAV עם טרופיזם עצבי. במחקר זה, מקדם ה- CBA שימש כמקדם חזק כדי לגרום לביטוי יתר של hαSyn, והסרוטיפ AAV5 שימש לווקטור הנגיפי. סרוטיפ זה שימש בעבר לתמרת נוירונים של עכברים וחולדות41,42. כאן, התוצאות הראו כי 12 שבועות לאחר מסירת AAV5-GFP ב-SN של עכברים, הפלואורסצנציה הירוקה הייתה נוכחת באופן סלקטיבי הן בצד האיפסילטרלי של ה-SN והן של הסטריאטום (איור 1), מה שמצביע על התמרה יעילה של נוירונים של המסלול הניגרוסטריאלי.
היבט קריטי נוסף של מודל העכבר הזה של מחלת פרקינסון הוא נקודת הזמן הנדרשת לניתוח תהליך מסוים לאחר הניתוח. בהקשר זה, עבודה זו מראה מחקר קינטי של תהליכים שונים המעורבים בפתולוגיה. מאחר שנקודות זמן מרכזיות משתנות עם המינון של הגנומים הנגיפיים הניתנים לכל עכבר, הסרוטיפ של AAV שבו נעשה שימוש, או אפילו עם אצווה של AAV בשימוש, בוצע תחילה ניתוח מינון-תגובה של כמות ה-AAV-αSyn הנדרשת כדי לגרום לאובדן משמעותי של נוירוני TH+ ולפגיעה מוטורית. מחקרים קודמים הראו פגיעה מוטורית משמעותית ואובדן נוירוני TH+ של המסלול הניגרוסטריאלי לאחר 12 שבועות של זריקות AAV-αSyn בעכברים במינונים הנעים בין 6 x 108-3 x 1010 גנומים ויראליים לכל עכבר 16,17,30,31. בהתאם לכך, המינון של AAV-hαSyn ששימש להשראת ביטוי hαSyn במסלול הניגרוסטריאלי, אובדן נוירוני TH+ ופגיעה מוטורית בעכברים נעו בין 1 x 108-1 x 1010 10 גנומים נגיפיים לכל עכבר. יתר על כן, כדי לשלוט בכך שאובדן נוירוני TH+ ופגיעה מוטורית הושרו על ידי ביטוי יתר של hαSyn ב- SN ולא על ידי זיהום AAV של נוירונים של SN, נכללו קבוצות ביקורת שבהן קידוד AAV עבור גן מדווח (AAV-eGFP) נמסר באופן חד צדדי ב- SN של עכברים ונקבעו ניוון עצבי ופגיעה מוטורית. התוצאות הראו ש-12 שבועות לאחר הניתוח הסטריאוטקסי, 1 x 1010 גנומים נגיפיים לכל עכבר היו מינון תקין של AAV5-hαSyn, שכן עכברים שקיבלו את העומס הנגיפי הזה הראו hαSyn משמעותי במסלול הניגרוסטריאטלי (איור 2 ואיור 3), אובדן נוירוני TH+ (איור 4) ופגיעה מוטורית (איור 5). לעומת זאת, מינונים נמוכים יותר של AAV5-hαSyn (1 x 108 גנומים נגיפיים לעכבר ו-1 x 109 גנומים נגיפיים לכל עכבר) לא היו חזקים מספיק כדי להגיע לשינויים משמעותיים בכל הפרמטרים האלה יחד (איורים 2-4). יש לציין כי מתן AAV-GFP ב-1 x 1010 10 גנומים נגיפיים לכל עכבר גרם לרמה נמוכה (כ-20%), אך משמעותית של אובדן תאי עצב מסוג TH+ של נוירונים דופמינרגיים ניגריים (איור 4A,B). תוצאה זו מסכימה עם תצפיות קודמות שהשתמשו במודל זה41 והיא ככל הנראה תוצאה של רמה נמוכה של דלקת עצבית המושרה על ידי מתן וקטורי AAV ב- SN. אף על פי כן, היקף האובדן של תאי עצב מסוג TH+ היה גבוה משמעותית בעכברים שקיבלו AAV5-hαSyn בהשוואה לאלה שקיבלו את אותו מינון של AAV-GFP (איור 4C). יש לציין כי הקינטיקה של ביטוי hαSyn תלויה לא רק ביעילות ההעתקה אלא גם בהיקף הדיפוזיה של AAV39. מאחר שדיפוזיית AAV תלויה בסרוטיפ AAV, נקודות הזמן המדויקות של המפתח במודל חייתי זה עשויות להשתנות בעת שימוש בסרוטיפ AAV אחר השונה מ-AAV5.
לאחר מכן, נערך ניתוח קינטי באמצעות 1 x 1010 10 גנומים נגיפיים לכל עכבר כדי לקבוע נקודות זמן מרכזיות במודל העכבר הזה. מכיוון שהראיות הנוכחיות הראו כמה תסמינים מוקדמים המופיעים לפני פגיעה מוטורית, אשר יאפשרו אבחון מוקדם של מחלת פרקינסון43,44, ניסויים אלה ביקשו למצוא את נקודת הזמן שבה ביטוי hαSyn כבר ניכר אך בהיעדר פגיעה מוטורית. התוצאות מראות כי הופעת הביטוי hαSyn ב-SN הייתה ב-5 שבועות לאחר המסירה הסטריאוטקסית של AAV-hαSyn (איור 6). נקודת זמן זו מהווה נקודה טמפורלית מעניינת להתחיל במתן טיפולים המותאמים לעצירת התהליכים הנוירו-דלקתיים והנוירודגנרטיביים. נקודות זמן מרכזיות נוספות שנקבעו כאן היו זמני השיא של שני אירועים קריטיים הקשורים לתהליך הדלקת העצבית: הזמן שבו המיקרוגליה מגיעה לדרגת ההפעלה המרבית וזמן החדירה המקסימלית של תאי T לתוך ה-SN. התוצאות הראו עקומה עם מגמה המגיעה לשני גלים של הפעלה מיקרוגליאלית מקסימלית, הראשון ב-10 שבועות לאחר הניתוח והשני ב-15 שבועות לאחר הניתוח (איור 7). הניתוח הקינטי של חדירת תאי T הראה את זמן השיא של חדירת Treg ל-SN ב-11 שבועות לאחר הניתוח הסטריאוטקסי (איור 8). באופן מפתיע, לא זוהו תאי T משפיעים (CD4+ Foxp3-) שחודרים ל-SN במהלך מסגרת הזמן שנותחה (שבועות 8-13 לאחר הניתוח). בסך הכל, תוצאות אלה מצביעות על פרק זמן מתאים להתחלת מתן טיפולים המיועדים לעצירת תהליך הדלקת העצבית והחלשת חדירת תאי T ל-SN באמצעות מודל פרה-קליני זה, שנע בין שבוע 5 לאחר הניתוח (תחילת ביטוי יתר של hαSyn) לבין שבוע 10 לאחר הניתוח (הגל הראשון של דלקת עצבית והסתננות תאי T) (איור 9).
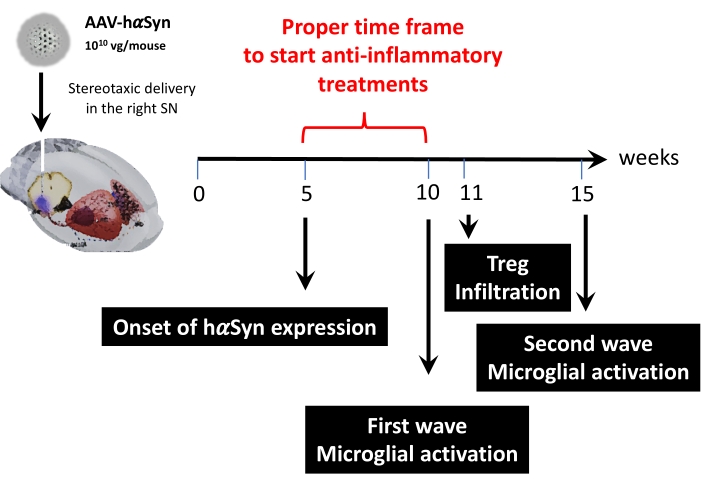
איור 9: סיכום של נקודות הזמן העיקריות שנמצאו עבור מודל זה של בעלי חיים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
