Al entregar y eliminar proteínas hacia y desde la membrana plasmática, el tráfico de proteínas de membrana en las neuronas controla el equipo de membrana plasmática con receptores y canales iónicos y, como resultado, regula la función neuronal. La mala regulación o los defectos en el tráfico de proteínas suelen tener efectos perjudiciales en las células y provocar una degeneración neuronal. En los seres humanos, esto puede causar enfermedades neurodegenerativas como la enfermedad de Alzheimer y Parkinson o la retinosis pigmentaria1. Los fotorreceptores en el ojo compuesto de Drosophila melanogaster se han convertido en un sistema modelo in vivo para estudiar el tráfico de proteínas de membrana2. Esto no solo se debe a la versatilidad genética de Drosophila que permite exámenes genéticos efectivos, sino también a que todos los componentes esenciales de la membrana fotorreceptora absorbente de luz se caracterizan con gran detalle y se dispone de técnicas microscópicas eficientes que se pueden aplicar al ojo de mosca. Estas técnicas son el foco de este artículo.
En las células fotorreceptoras de Drosophila, la membrana plasmática apical forma una pila densamente empaquetada de microvellosidades a lo largo de un lado de la célula, denominada rabdomero. Los rabdomeros de las células fotorreceptoras R1-6 están dispuestos en un patrón trapezoidal característico, mientras que las células fotorreceptoras R7 y R8 forman un solo rabdomero en el centro de este trapecio3. El tráfico de proteínas de membrana es necesario para un recambio regulado de proteínas de membrana rabdomeral como la rodopsina y los canales iónicos TRP (potencial de receptor transitorio) y TRPL (similar a TRP) activados por la luz para asegurar la cantidad adecuada de estas proteínas de fototransducción en el rabdomero. Las proteínas de membrana fotorreceptoras se sintetizan en el retículo endoplásmico y se transportan a través del aparato de Golgi al rabdomero. Después de la activación de la rodopsina por la luz, una molécula de rodopsina puede inactivarse por absorción de un segundo fotón o puede ser eliminada del rabdomero por endocitosis mediada por clatrina. La rodopsina endocitosa se degrada en el lisosoma o se recicla de nuevo al rabdomero 4,5. El canal iónico TRPL también se internaliza tras la activación de la cascada de fototransducción y sufre una translocación dependiente de la luz entre el rabdomero (donde se encuentra cuando las moscas se mantienen en la oscuridad) y un compartimento de almacenamiento enriquecido con ER en el cuerpo celular (al que se transporta dentro de varias horas después de la iluminación)6,7,8,9,10 . A diferencia de la rodopsina endocitosa, solo pequeñas cantidades de TRPL se degradan a través de la vía endolisosomal, y la mayoría se almacena intracelularmente en su lugar y se recicla de nuevo al rabdomero tras la adaptación oscura6. Por lo tanto, TRPL se puede utilizar para analizar el tráfico de proteínas de membrana plasmática desencadenado por la luz. Las células fotorreceptoras de Drosophila también se emplean para estudiar la degeneración neuronal. La degeneración de las células fotorreceptoras se determina con frecuencia mediante la evaluación de la estructura de los rabdomeros, que se desintegran como resultado de procesos degenerativos5.
Con el fin de estudiar la localización subcelular de TRPL y rodopsina en células fotorreceptoras o degeneración de células fotorreceptoras, se han aplicado aquí dos métodos de microscopía de fluorescencia que difieren con respecto a la velocidad de análisis y la resolución. Un método muy rápido y no invasivo que se puede utilizar para cribados genéticos pero con una resolución espacial limitada es la detección de fluorescencia en el pseudopúpilo profundo (DPP). El DPP es un fenómeno óptico de ojos compuestos de artrópodos cuyo origen geométrico ha sido explicado en detalle por Franceschini y Kirschfeld en 197111. En resumen, en varios planos ópticos debajo de la superposición de retina se pueden observar imágenes de rabdomeros de ommatidios adyacentes. En un plano focal a través del centro de la curvatura del ojo, estas proyecciones superpuestas forman una imagen que se asemeja al diseño trapezoidal de rabdomeros en un solo ommatidio solo órdenes de magnitud más grande. Este fenómeno también se puede observar independientemente de la expresión exógena de proteínas de fluorescencia (por ejemplo, TRPL::eGFP8), que sin embargo hacen que el DPP sea más fácil de detectar (Figura 1A-A”)12. Un segundo método no invasivo es la microscopía de inmersión en agua que se basa en la obtención de imágenes de proteínas marcadas con fluorescencia después de neutralizar ópticamente el aparato dióptrico de los ojos con agua (Figura 1B-C”)12. Utilizando el método de inmersión en agua, la cantidad relativa de TRPL::eGFP en los rabdomeros o cuerpo celular se puede evaluar cuantitativamente para células fotorreceptoras individuales. Además, las proteínas marcadas con fluorescencia no translocadoras se pueden utilizar para evaluar la integridad rabdomeral y determinar el curso temporal de una degeneración potencial de manera cuantitativa, como se describe aquí.
Si bien las grabaciones del DPP son, con mucho, los métodos más fáciles y rápidos de realizar, la resolución espacial de los datos que generan es limitada. Además, hay numerosas razones por las que un DPP puede estar ausente, que no son necesariamente discernibles por las propias imágenes de DPP. Dado que el DPP representa una suma de varios ommatidios, se pierde información sobre células individuales. Por lo tanto, las imágenes DPP de baja resolución cumplen una función importante en la detección de un gran número de moscas, pero generalmente deben ir seguidas de grabaciones de mayor resolución a través de microscopía de inmersión en agua. Las micrografías de inmersión en agua permiten interpretaciones sobre células individuales, defectos del desarrollo, morfología ocular, mala localización de proteínas o degeneración de la retina, así como la cuantificación de estos efectos. Este Protocolo describe estas dos técnicas en detalle.
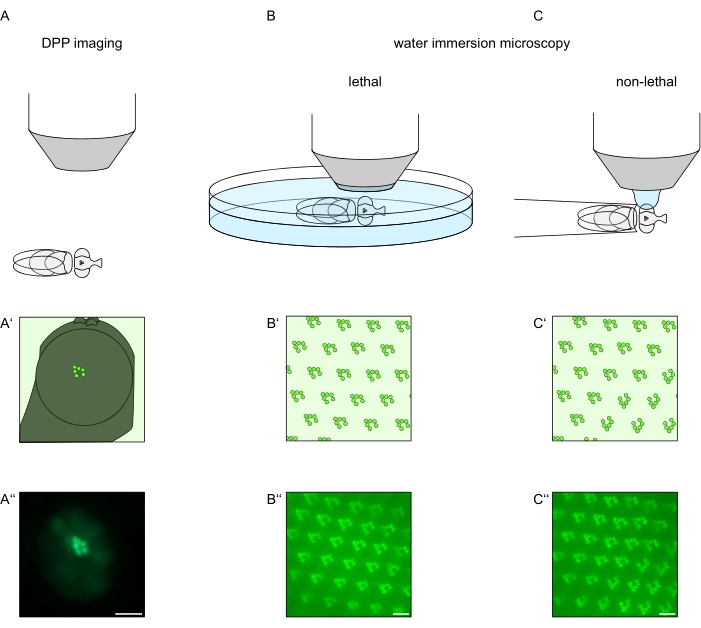
Figura 1: Descripción general de las variaciones de microscopía para el ojo de Drosophila presentadas en este Protocolo. Representaciones esquemáticas y micrografías ejemplares de imágenes de pseudopúpodo profundo fluorescente (DPP) (A-A‘‘), microscopía letal de inmersión en agua (B-B”) de rabdomeros fluorescentes y (C-C”) microscopía de gota de agua no letal de rabdomeros fluorescentes. Barra de escala (A”): 100 μm. Barras de escala (B”–C”): 10 μm. La figura ha sido modificada a partir de la referencia13. Haga clic aquí para ver una versión más grande de esta figura.
