Senescenta celler ackumuleras normalt i organismer under år under normalt biologiskt åldrande men kan också utvecklas snabbt i tumörceller som ett svar på skador som induceras av olika cancerbehandlingar, inklusive strålning och kemoterapi. Även om de inte längre sprider sig kan terapiinducerade senescenta (TIS) tumörceller bidra till behandlingsresistens och driva återfall 1,2,3. Faktorer som utsöndras av TIS-celler kan förvärra tumörmalignitet genom att främja immunflykt eller metastasering 4,5. TIS-celler utvecklar komplexa, kontextspecifika fenotyper, förändrade metaboliska profiler och unika immunsvar 6,7,8. Därför är identifiering och karakterisering av TIS-tumörceller inducerade av olika cancerbehandlingsmetoder ett ämne av pågående intresse för cancerforskningssamhället.
För att detektera TIS-tumörceller används konventionella åldrandeanalyser i stor utsträckning, främst baserat på att detektera ökad aktivitet hos åldrandemarkörenzymet, den lysosomala beta-galaktosidasen GLB19. Detektion vid ett nästan neutralt (snarare än surt) lysosomalt pH möjliggör specifik detektion av åldrande-associerat beta-galaktosidas (SA-β-Gal)10. En standard SA-β-Gal-analys som har använts i flera decennier använder X-Gal (5-brom-4-klor-3-indolyl-β-D-galaktopyranosid), ett blått kromogent beta-galaktosidassubstrat, för att detektera SA-β-Gal i fasta celler med ljusmikroskopi11. X-Gal-analysen möjliggör kvalitativ visuell bekräftelse av TIS med hjälp av allmänt tillgängliga reagenser och laboratorieutrustning. Ett grundläggande överfört ljusmikroskop är den enda instrumentering som krävs för att utvärdera närvaron av det blå kromogenet. X-Gal-färgningsproceduren kan dock sakna känslighet, vilket ibland kräver mer än 24 timmar för att färgen ska utvecklas. Färgning följs av låg genomströmning, subjektiv poängsättning av enskilda senescenta celler baserat på att räkna cellerna som uppvisar en viss intensitetsnivå av den blå kromogenen under ett ljusmikroskop. Eftersom X-Gal är cellogenomtränglig kräver denna analys lösningsmedelsfixerade celler, som inte kan återvinnas för nedströmsanalys. När man arbetar med begränsade prover från djur eller patienter kan detta vara en stor nackdel.
Förbättrade SA-β-Gal-analyser med cellpermeant, fluorescerande enzymsubstrat, inklusive C 12-FDG (5-dodecanoylaminofluorescein Di-β-D-Galactopyranoside, grön) och DDAOG (9H-(1,3-diklor-9,9-dimetylakridin-2-en-7-yl) β-D-Galactopyranoside, långt röd) har tidigare förekommit i litteraturen12,13,14,15. DDAOG:s kemiska sondstruktur och optiska egenskaper visas i kompletterande figur S1. Dessa cellpermeanta sonder möjliggör analys av levande (snarare än fasta) celler, och fluorescerande snarare än kromogena sonder underlättar användningen av snabba fluorescerande analysplattformar med hög genomströmning, inklusive screeninginstrument med högt innehåll och flödescytometrar. Sorteringsflödescytometrar möjliggör återhämtning av berikade populationer av levande åldrande celler från cellkulturer eller tumörer för nedströmsanalys (t.ex. western blotting, ELISA eller ‘omics). Fluorescensanalys ger också en kvantitativ signal, vilket möjliggör mer exakt bestämning av andelen åldrande celler i ett givet prov. Ytterligare fluorescerande sonder, inklusive viabilitetssonder och fluoroformärkta antikroppar, kan lätt läggas till för multiplexerad analys av mål utöver SA-β-Gal.
I likhet med DDAOG är C12-FDG en fluorescerande sond för SA-β-Gal, men dess gröna fluorescerande utsläpp överlappar med inneboende cellulär AF, som uppstår under åldrande på grund av ackumulering av lipofuscinaggregat i celler16. Genom att använda den fjärrröda DDAOG-sonden kan grön cellulär AF användas som en sekundär parameter för att bekräfta åldrande17. Detta förbättrar analysens tillförlitlighet genom att använda en andra markör utöver SA-β-Gal, som ofta kan vara opålitlig som en enda markör för åldrande18. Eftersom detektion av endogen AF i åldrande celler är ett etikettfritt tillvägagångssätt är det ett snabbt och enkelt sätt att utöka specificiteten hos vår DDAOG-baserade analys.
I detta protokoll demonstrerar vi användningen av DDAOG och AF som en snabb, dubbelparameterflödescytometrianalys för identifiering av livskraftiga TIS-tumörceller från in vitro-kulturer eller isolerade från läkemedelsbehandlade tumörer etablerade hos möss (Figur 1). Protokollet använder fluoroforer som är kompatibla med ett brett spektrum av vanliga kommersiella flödescytometrianalysatorer och sorterare (tabell 1). Kvantifiering av procentandelen livskraftiga senescenta celler med hjälp av standardflödescytometrianalys är aktiverad. Om så önskas kan ett valfritt immunmärkningssteg utföras för att utvärdera cellyteantigener av intresse samtidigt med åldrande. Identifierade senescenta celler kan också berikas med hjälp av standard fluorescensaktiverad cellsorteringsmetodik (FACS).
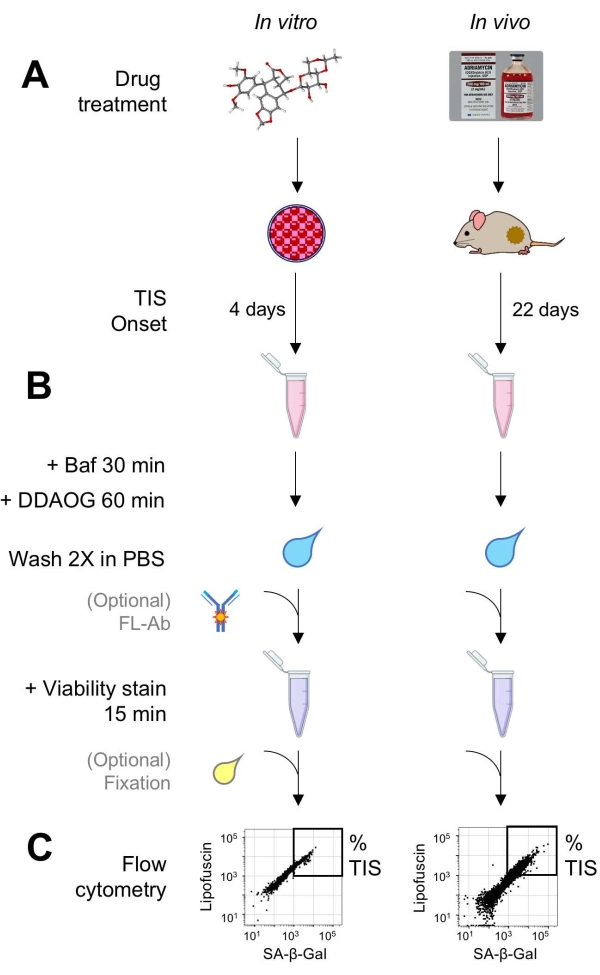
Bild 1: Experimentellt arbetsflöde. Ett schema som sammanfattar viktiga punkter i DDAOG-analysen. (A) Ett TIS-inducerande läkemedel tillsätts till däggdjursodlade celler eller administreras till tumörbärande möss. Tid tillåts sedan för uppkomsten av TIS: för celler, 4 dagar efter behandling; för möss, totalt 22 dagar, med tre behandlingar var 5: e dag plus 7 dagars återhämtning. Celler skördas eller tumörer dissocieras i suspension. B) Proverna behandlas med Baf för att justera lysosomalt pH för detektion av SA-β-Gal i 30 minuter. Sedan läggs DDAOG-sonden till i 60 minuter för att detektera SA-β-Gal. Proverna tvättas 2x i PBS, och en viabilitetsfläck tillsätts kort (15 min). Eventuellt kan prover färgas med fluorescerande antikroppar i öppna fluorescenskanaler och/eller fixeras för senare analys. (C) Proverna analyseras med hjälp av en standardflödescytometer. Livskraftiga celler visualiseras i punktdiagram som visar röd DDAOG (indikerar SA-β-Gal) kontra grön autofluorescens (lipofuscin). En grind för att bestämma procentandelen TIS-celler upprättas baserat på obehandlade kontrollprover (visas inte). Om en sorteringscytometer (FACS) används kan TIS-celler samlas in och placeras tillbaka i odling för ytterligare in vitro-analyser eller lyseras och bearbetas för molekylärbiologiska analyser. Förkortningar: DDAO = 9H-(1,3-diklor-9,9-dimetylakridin-2-on); DDAOG = DDAO-Galaktosid; TIS = terapiinducerad åldrande; FL-Ab = fluoroforkonjugerad antikropp; Baf = Bafilomycin A1; SA-β-Gal = åldrande-associerad beta-galaktosidas; PBS = fosfatbuffrad saltlösning; FACS = fluorescensaktiverad cellsortering. Klicka här för att se en större version av denna siffra.
| Fluorofor | Upptäcker | Ex/Em (nm) | Cytometerlaser (nm) | Cytometerdetektor / bandpassfilter (nm) |
| DDAOG | SA-β-Gal | 645/6601 | 640 | 670 / 30 |
| AF | Lipofuscin | < 600 | 488 | 525 / 50 |
| CV450 | Livsduglighet | 408/450 | 405 | 450 / 50 |
| PE | Antikropps-/ytmarkör | 565/578 | 561 | 582 / 15 |
Tabell 1: Optiska specifikationer för fluoroforer och cytometer. Cytometerspecifikationer som används i detta protokoll listas för ett instrument med totalt 4 lasrar och 15 emissionsdetektorer. DDAOG detekterad vid 645/660 nm är formen av sonden klyvd av SA-β-Gal1. Uncleaved DDAOG kan uppvisa låg fluorescens vid 460/610 nm men avlägsnas genom tvättsteg i protokollet. Förkortningar: DDAO = 9H-(1,3-diklor-9,9-dimetylakridin-2-on); DDAOG = DDAO-Galaktosid; AF = autofluorescens; PE = phycoerythrin; SA-β-Gal = åldrande-associerad beta-galaktosidas.
