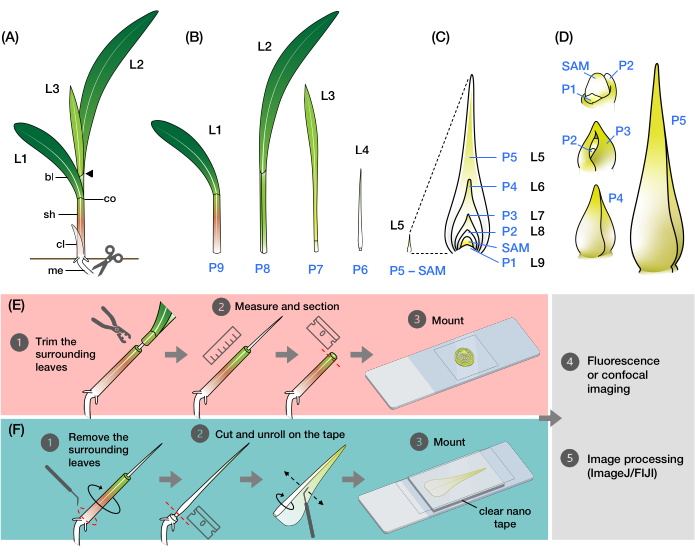
1. Iscensättning av majsbladutveckling och utformning av experimentet Växande växter och iscensättning av bladutvecklingBestäm ålder och utvecklingsstadium för de växter som ska användas för experimentet och utför en detaljerad iscensättning av bladutveckling.OBS: Det rekommenderas att iscensättningen utförs på växter med samma genetiska bakgrund som de muterade eller transgena linjer som kommer att användas i experimenten. SAM slutar producera nya blad mellan 20 och 25 DAG, beroende på den genetiska bakgrunden och tillväxtförhållandena. Av denna anledning är de metoder som beskrivs här idealiska för majsplantor i åldern 7-14 DAG. Odla majsplantorna i ett växthus eller en tillväxtkammare enligt ett lämpligt växtskyddsprotokoll35. Samla växterna i önskad ålder eller V-steg med en liten kniv eller sax genom att skära vid mesocotyl, som är stammen under markytan (figur 1A). Sätt försiktigt in skärverktyget i jorden bredvid plantan och skjut ner det till basen i 45 ° vinkel för att skära mesocotylen. Lyft plantan ur jorden och damma bort eventuell kvarvarande smuts eller jordpartiklar som kan klamra sig fast vid växten. Ta bort coleoptile, den skyddande manteln som täcker den framväxande skottet i unga plantor, för hand. För varje växt, registrera V-steget, antalet blad och längden på det sista bladet som kommer från virveln. Ta bilder av växterna för framtida referens. Ta bort de äldre bladen en efter en. För att göra detta, håll växten genom resterna av mesocotyl eller stammen och skära ut varje blad från mantelns botten och veckla försiktigt ut manteln på ett cirklande sätt med en tandsond (figur 1B).Se steg 2.1 för en snabb metod för att ta bort de omgivande bladen. Under ett stereomikroskop med cirka 2x förstoring, dissekera försiktigt resten av skottspetsen på fuktiga våtservetter för att förhindra att provet torkar ut. Punktbeskatta försiktigt och veckla ut varje bladprimordium med en tandsond med en böjd nål eller ett par fina pincett tills SAM, P1 och P2 är synliga (figur 1D). Registrera utseende och mätning av varje plastokron.OBS: Se ett exempel på bladutveckling i majsplantor i kompletterande fil 1. Planera experimentetOBS: Att planera experimentet noggrant kan förbättra effektiviteten. Till exempel, i tvärsnittsanalys, kan bestämning av den ungefärliga platsen som ska skäras vid avbildning av specifika plastokroner eller regioner minska dissektionstiden. Figur 2 visar ett exempel på standardiserad provtagning. Dessutom kan optimering av bildparametrarna baserat på egenskaperna hos FP eller fluorescerande sonder förbättra experimentens effektivitet.Använd iscensättningsinformationen som en guide för att fokusera experimentet på specifika plastokroner, primordiumregioner och vävnader. Använd FP eller andra fluorescerande sonder av intresse för att följa avsnitt 2 och / eller 3 nedan på några prover.Anmärkning: Se tillgängliga utsädeslager för majs FP linje 36-42 vid https://www.maizegdb.org/data_center/stock. Bestäm de optimala bildinsamlingsparametrarna för varje FP-reporter eller sond, inklusive lämplig detektor eller filter, excitations- och emissionsvåglängder, pinhålstorlek och andra inställningar. Spara inställningarna för att återskapa arbetsförhållandena för vart och ett av experimenten.OBS: Se Linh och Scarpella30 för några tekniska riktlinjer för konfokal laserskanningsmikroskopi av framväxande löv. Planera bildtagningstiden baserat på halveringstiden och andra egenskaper hos FP-reportern43.OBS: Tiden mellan dissektion och avbildning måste också beaktas vid planering av ett experiment, eftersom färska växtprover utsätts för uttorkning och cellförändringar under denna period. Figur 2: Provtagningsschema för tvärsnittsanalys av primordia av majsblad. (A, vänster) Proximalt skott av en 7 DAG-majsplanta som visar ett exponerat fjärde blad (L4) vid P7. De brutna linjerna anger de sju provtagningspunkterna längs primordiet, från 0,5 mm till 10 mm. (A, höger) Schematisk representation av bladprimordia, med den projicerade storleken och positionen för varje plastokron: P7 (vit); P6 (magenta); P5 (blå); P4 (grön); och P3 tills SAM (gul). (B-H) Fluorescensbilder av tvärsnitt som representerar provtagningspunkterna i A från 10 mm (B) till 0,5 mm (H). Primordia är pseudofärgade enligt plastochronfärgschemat i (A). Sektionerna avbildades med ett epifluorescensmikroskop med hjälp av ett UV-filter för långpassemission för autofluorescens. Skalstång = 200 μm (B-H). Denna figur har modifierats och reproducerats med tillstånd från Robil och McSteen24. Förkortningar: DAG = dagar efter spiring; P = plastokron; SAM = skjuta apikal meristem; † = bladmantel eller egentlig förligula, * = skjut apikalt meristem; ** = stam. Klicka här för att se en större version av denna figur. 2. Avbildning av tvärsnitt av majsblad primordia Strippa äldre omgivande löv och mäta bladprimordiumSamla upp plantan genom att försiktigt klippa den vid mesocotylen under markytan med en liten kniv eller sax, enligt beskrivningen i steg 1.1.3-1.1.4 (figur 1A). Bestäm rätt trådborttagningshålstorlek som ska användas och var snittet ska göras längs skottet. Se till att hålet är tillräckligt stort för att passa målet primordium. Börja klippa vid den mer distala delen av skottet och arbeta gradvis mot basen tills primordiumets spets exponeras. För att göra snittet, håll trådstripparen med käftarna mot skottet. Placera skottet mot det valda hålet och pressa ihop stripparhandtagen för att skära igenom bladvirveln.OBS: För att undvika att oavsiktligt skära målprimordiet, rikta försiktigt in skottets mitt med hålet i trådstripparen. För större växter rekommenderas att manuellt ta bort några av de omgivande bladen för hand innan du använder stripparen.VARNING: Trådstrippare har vassa käkar som kan orsaka skärsår eller andra skador om de inte används försiktigt. Medan du fortsätter att pressa ihop handtagen, skjut stripparen bort från skottet för att trimma den övre delen av bladen. Se till att målområdet i primordium förblir täckt av de omgivande bladen (kompletterande figur S1C, D).OBS: Efter trimning av de övre bladen lämnas skottet med en exponerad primordiumspets (P5, P6 eller P7, beroende på snittets position och storleken på det trådstripphål som används; Figur 1E). Detta primordium kommer att fungera som referens för att uppskatta storleken och stadiet av yngre primordia. P6 är den mest praktiska referensen att använda i majsplantor på grund av dess storleksintervall och stiftliknande morfologi. Använd en linjal för att mäta längden från primordiumets spets till skottets botten. Registrera mätningen.OBS: Längden på primordium kommer att överskattas något eftersom det finns några millimeter stam vid basen av primordia (figur 2A). Frihandssnittning och avbildning av bladprimordiumUnder ett stereomikroskop med cirka 0,8x förstoring, håll skottet stadigt på en slät yta, till exempel en skärbräda av glas eller annat reptåligt material. Använd ett rakblad eller skalpell för att skära tunna sektioner (0,25-0,8 mm) av skottet vid önskade punkter längs primordiumets längd. Gör ett första snitt något ovanför målområdet för att kasta den distala delen av skottet och få sedan tunna sektioner runt målområdet. För att göra rena snitt, håll bladet vinkelrätt mot skottet och skjut det inåt med en jämn, jämn rörelse.OBS: Det kan finnas några millimeter sektionsutrymme för större primordia (P6 eller P7). För mindre primordia (P5 och tidigare) kan dock 1 mm göra en signifikant skillnad i provtagningen, eftersom dessa primordier staplas upp inom de första millimeterna från skottets bas (se figur 2A, F-H). Det rekommenderas att använda ett tveeggat rakblad för finare sektionering.VARNING: Var försiktig när du använder rakblad och skalpeller för att undvika oavsiktliga skärsår eller skador. Byt ut rakbladet regelbundet, eftersom bladmantlarna lätt kan slöa bladet. Täck skottet med fuktiga våtservetter efter varje sektionsomgång för att förhindra att provet torkar ut. Montera bladsektionen på en ren glasskiva (75 mm x 25 mm), använd en pipett för att applicera en droppe vatten (eller 50% glycerol) direkt på sektionen och placera en täckglas (22 mm x 22 mm) ovanpå. Om du använder ett fluorescerande färgämne, applicera färglösningen på sektionen, placera en täckglas ovanpå och inkubera den efter behov.OBS: Beroende på typen av färgämne kan ytterligare steg för bearbetning av bladsektionerna vara nödvändiga innan du fortsätter till nästa steg. Dessa referenser44,45,46,47 ger allmänna detaljer och användningar av fluorescerande färgämnen vid avbildning av växtceller. Ett kärnfärgande fluorescerande färgämne, 5-etynyl-2′-deoxiuridin (EdU), har använts effektivt på tvärsnitt av majsblad primordia48. Däremot ger plasmamembranfärgningsfärgämnen, såsom Fei Mao 4-64 (FM 4-64) och propidiumjodid, inte tillfredsställande resultat (figur 3A-D). Placera bilden på scenen i ett epifluorescens- eller konfokallaserskanningsmikroskop och justera fokus och inställningar efter behov för att visualisera fluoroforen30. Se tabell 1 för de belysnings- och bildinsamlingsinställningar som används för utvalda FP-reporterlinjer för majs.OBS: Tjocka sektioner kan ge höga nivåer av bakgrundsautofluorescens. Överväg några strategier för bildbehandling och provberedning som kan hjälpa till att mildra detta problem30,49 (se tabell 2).VARNING: Var försiktig och använd skyddsglasögon när du arbetar med kraftfulla ljuskällor och lasrar för att minska risken för ögonskador. Bild genom bladsektionen vid olika förstoringar och detekteringskanaler (inklusive ljusfältsbelysning)30. Använd 4x förstoring för att avbilda hela tvärsnittet och 20x eller 40x förstoring för att avbilda specifika vävnader. Minska exponeringstiden vid högre förstoringar för epifluorescensavbildning för att skydda proverna från snabb fotoblekning (se tabell 1). Ta bilder av hela avsnittet eller specifika intressanta regioner (ROI). Om det behövs, skaffa en z-stack, som är en serie bilder som tagits med olika brännvidder30. För att få en z-stack, bestäm först de övre och nedre positionerna i provet, sedan antalet optiska sektioner som måste fångas mellan dessa positioner. Färre optiska sektioner resulterar i en större z-stegstorlek, vilket är avståndet mellan varje sektion. Beroende på FP: s egenskaper och den förstoring som används, fånga tre till 25 optiska sektioner med z-stegstorlekar mellan 1 och 12 μm för avbildning av tvärsnitt av majsbladprimordia.OBS: För att platta z-stacken, använd en lämplig volymåtergivningsteknik50 som kan åstadkommas genom mikroskopprogramvaran eller genom att använda ImageJ / FIJI34 (se steg 4.1 och 4.2). Spara alla bildfiler och tillhörande metadata, om sådana finns. Tabell 1: Inställningar för belysning och bildinsamling som används för fluorescens och konfokal avbildning av utvalda rapportörer för majs. Förkortningar: FP = fluorescerande protein; TRITC = tetrametylrodamin; FITC = fluoresceinisotiocyanat, WLL = laser med vitt ljus; Ar-jon = argonjonlaser; HyD = hybriddetektor; AU = Luftig enhet; Hz = Hertz, skanningslinje per sekund. Klicka här för att ladda ner denna tabell. 3. Avbildning rullade ut hela fästen av majsblad primordia Dissekera primordia och förbereda glasskivan med nanotejpSamla upp plantan genom att försiktigt klippa den vid mesocotylen under markytan med en liten kniv eller en sax, enligt beskrivningen i steg 1.1.3-1.1.4 (figur 1A). Förbered glasskivan (75 mm x 25 mm) med den klara, dubbelhäftande nanotejpen (kompletterande figur S1H) innan du dissekerar växten. Klipp en rektangulär bit nanotejp och fäst den i mitten av en ren glasskiva. Ta inte bort den skyddande plastfilmen på tejpens ovansida.OBS: Bestäm storleken på tejpen som ska användas enligt storleken på proverna som ska avbildas (se exempel på överdimensionerade prover i kompletterande figur S1I, J). Nanotejpen finns i antingen akryl- eller polyuretangelmaterial. Båda typerna är effektiva för fluorescens och konfokal avbildning24. För att förhindra missfärgning av tejpen, skydda den från långvarig exponering för ljus genom att förvara den i en mörk behållare. Börja dissekera skottet genom att ta bort den övre delen av de äldre bladen med en trådstrippare, som beskrivs i steg 2.1. Håll skottet vid mesocotylen och ta försiktigt bort de omgivande bladen med en tandsond för att extrahera primordia. För att göra detta, ta bort bladen från skottets botten och veckla ut dem en i taget tills primordium av intresse exponeras. Alternativt kan du använda trådstripparen för att direkt extrahera bladprimordia genom att göra ett snitt vid skottets basala region (kompletterande figur S1E-G).OBS: Denna metod medför en högre risk att knäppa primordia. Montering och avbildning av primordium.Ta bort skyddsfilmen från tejpen. Lägg det exponerade primordium på tejpen. Använd ett rakblad för att skära primordium vid basen, kassera basalstammen och hypokotylen (figur 1F). Rulla ut primordium med en tandsond med en böjd nål (spetsdiameter på 0,25-0,6 mm). Använd en mikrosond (spetsdiameter på 0,15-0,2 mm) för primordia mindre än 3 mm (P3 till tidig P4). Placera nålspetsen parallellt med ytan och veckla ut den yttre basalmarginalen, tryck den försiktigt mot tejpen. För mjukare utrullning, smörj bladets inre yta (adaxial) genom att doppa nålspetsen i 100% glycerol. Med den yttre marginalen fäst vid tejpen, rikta in nålspetsen parallellt med bladets långa axel. Skjut försiktigt nålen i sidled för att rulla ut och platta bladet på tejpen (figur 1F).OBS: Att bryta eller skada bladet under rullning är vanligt, särskilt i större primordia med en robust midrib (se figur 3E-G). Att rulla ut bladet kraftigt kan blåsa ytan och producera artefakter under avbildning (figur 3H). Se tabell 2 och diskussionen för rekommenderade lösningar på dessa problem. Applicera en droppe vatten på det orullade primordiumet. Lägg omedelbart ett täckglas ovanpå vattendroppen och primordiumet. Tryck försiktigt ner kanterna på täckglaset så att de fäster vid tejpen.OBS: Det rekommenderas att använda stora rektangulära täckglas (50 mm x 24 mm eller 60 mm x 24 mm) med extra områden för att fästa på tejpen. Minimera bildandet av luftbubblor, vilket kan förvränga bladet och orsaka ojämn ljusbrytning (se figur 3I-K). Se till att täckglaset fäster helt på tejpen, eftersom marginalerna på det monterade primordium kan rulla tillbaka in under ett löst vidhäftat täckglas (se figur 3L). Placera bilden på scenen av en epifluorescens (se kompletterande figur S1K) eller konfokalt laserskanningsmikroskop och justera fokus och inställningar efter behov för att visualisera fluoroforen30. Se tabell 1 för de belysnings- och bildinsamlingsinställningar som används för de valda rapportlinjerna för majs. Avbilda hela bladet eller en specifik ROI genom olika detekteringskanaler30. För att avbilda hela bladet, antingen manuellt fånga mosaikbilder av bladet eller använd mikroskopets kakeloperation vid låg förstoring (4x eller 10x).OBS: Även vid den lägsta förstoringen kan majsbladprimordia vara för stor för att avbildas med ett epifluorescens- eller konfokallaserskanningsmikroskop, vilket resulterar i längre bildtider som kan resultera i fotoblekning (se figur 3M) eller ökad bakgrundsautofluorescens. Därför rekommenderas att man använder ett fluorescensstereomikroskop för avbildning av helbladsprover större än 5 mm. För konfokal avbildning är det nödvändigt att erhålla z-staplar som omfattar bladets tjocklek (se steg 2.2.9). Spara alla bildfiler och tillhörande metadata, om sådana finns. Tabell 2: Felsökning av vanliga problem vid avbildning av tvärsnitt och hela fästen av majsbladprimordia. Klicka här för att ladda ner denna tabell. Figur 3: Suboptimal tvärsnitt och helmonterad beredning av majsbladprimordia. (A-D) Representativa konfokala bilder av tvärsnitt av bladprimordia med en plasmamembranmarkör, PIP2-1-CFP (A), och ett plasmamembranbindande fluorescerande färgämne, FM 4-64 (B), med motsvarande ljusfältsbilder (C,D). Jämfört med PIP2-1-CFP visar FM 4-64 suboptimal visualisering av cellkonturer. (E-M) Representativa fluorescensbilder av hela fästen av bladprimordia som visar förekomst av rivning (E-G), blåslagna ytor† (H), luftbubblor* (I–K), bakåtrullade marginaler (L) och fotoblekta områden‡ (M). Bladprimordia uttrycker DII-Venus (E-G), GAR2-YFP (H-J), mDII-Venus (K), PIN1a-YFP (L) och DR5-RFP (M). Skalstapel = 200 μm (AD); 500 μm (E-M). Figur 3A har modifierats och reproducerats med tillstånd från Robil och McSteen24, medan figur 5B-M är opublicerade data från författarna. Förkortningar: P = plastokron; YFP = gult fluorescerande protein; RFP = rött fluorescerande protein; CFP = fluorescerande cyanprotein, PIP2-1-CFP = pZmPIP2-1::ZmPIP2-1:CFP; DII-Venus = pZmUbi:DII:YFP-NLS; GAR2-YFP = p Zm GAR2::ZmGAR2:YFP; mDII-Venus = pZmUbi:mDII:YFP-NLS; PIN1a-YFP = p Zm PIN1a::ZmPIN1a:YFP; DR5-RFP = DR5rev::mRFPer; BF = ljusfält. Klicka här för att se en större version av denna figur. 4. Bearbetar bilderna med ImageJ/FIJI Platta z-staplar med maximal intensitetsprojektion (MIP)Starta ImageJ/FIJI och öppna stapeln med bilder, eller skapa en ny stapel från flera separata avbildningar genom att välja Image | Travar | Bilder att stapla från menyn. Välj bildstapeln. På menyn väljer du Bild | Travar | Z-projektet…. I dialogrutan Z-Project väljer du Max intensitet i listrutan Projektionstyp . Klicka på OK för att skapa maximal intensitet z-projektion.OBS: Den platta bilden är en 2D-projektion av pixlarna med den högsta intensiteten över z-stacks50. Denna metod är lämplig för att visualisera fluoroforen men inte de anatomiska egenskaperna i ljusfältsbilder av tjocka tvärsnitt och helbladsfästen. Spara bilden som TIFF (Tagged Image File Format) genom att välja Arkiv | Spara som | Tiff. Ange ett namn för filen och klicka på Spara.OBS: Det rekommenderas att använda TIFF för att lagra fluorescens och konfokala bilder, eftersom det behåller kvaliteten och detaljerna när bilden sparas. Platta z-staplar med hjälp av EDF-plugin (Extended Depth of Field)51Installera EDF-plugin52. Ladda ner Extended_Depth_Field.jar-filen (version 17.05.2021) från http://bigwww.epfl.ch/demo/edf/ och placera den i mappen “plugins” i ImageJ/FIJI. Starta ImageJ/FIJI och öppna stapeln med bilder, eller skapa en ny stapel från flera separata avbildningar genom att välja Image | Travar | Bilder att stapla från menyn. Välj bildstapeln. På menyn väljer du Plugins | Utökat skärpedjup | EDF Enkelt läge. I dialogrutan Utökat skärpedjup använder du skjutreglagen under Hastighet/ kvalitet för att ställa in önskad kvalitet och bearbetningshastighet och reg för höjdkarta för att ange jämnhetsnivå för topografin. Klicka på Kör för att skapa den rekonstruerade avbildningen.EDF kombinerar stacken till en skarp, sammansatt 2D-bild genom att projicera bästa fokus51. Denna teknik är användbar för att övervinna det begränsade fokusdjupet och är lämplig för att rekonstruera ljusfältsbilder av tjocka tvärsnitt eller helbladsfästen. Var försiktig när du använder det här filtret för att visualisera fluoroforen eftersom EDF, till skillnad från MIP, justerar bildens kontrast, vilket påverkar pixelintensiteten. Spara bilden som en TIFF-fil genom att välja Arkiv | Spara som | Tiff. Ange ett namn för filen och klicka på Spara. Sammanfoga bilder med plugin-programmet Grid/Collection Stitching53Spara samlingen av bilder eller bildstaplar som ska sys ihop i en enda katalog. Starta ImageJ/FIJI och välj Plugins | Sömmar | Rutnät/samling sömmar från menyn. I dialogrutan Sammanfogning av rutnät/samling väljer du Okänd position i listrutan Typ och sedan Alla filer i katalogen från Ordning. Klicka på OK. I dialogrutan Rutnätssömmar: Okänd position, Alla filer i katalogen anger du platsen för bilderna genom att skriva eller klistra in katalogsökvägen i textfältet Katalog eller genom att klicka på Bläddra… för att hitta katalogen. Klicka på OK för att starta sömnadsprocessen.Alternativet Okänd position i plugin-programmet Grid/Collection Stitching rekonstruerar en sammansatt bild från en uppsättning sammanhängande bilder genom att analysera deras överlappande regioner. Därför är det här alternativet användbart för att sy mosaik av bilder av tvärsnitt eller helbladsfästen som inte erhålls med hjälp av en kakelskanning. När sömnaden är klar visas ett nytt fönster med den sydda bilden. Spara bilden som en TIFF-fil genom att välja Arkiv | Spara som | Tiff. Ange ett namn för filen och klicka på Spara.OBS: Sammanfogning av stora montage av 2D- och 3D-bilder kan också åstadkommas med hjälp av kommandon för bildrekonstruktion i mikroskopprogramvaran eller genom importalternativen för bioformat i ImageJ/FIJI. Kombinera flera kanaler med kommandot Sammanfoga kanalerStarta ImageJ / FIJI och öppna bilderna eller bildstaplarna som ska slås samman.Bildstaplar kan först plattas ut med steg 4.1 eller 4.2. MIP rekommenderas generellt för att platta ut fluoroforkanaler, medan EDF är bättre lämpad för att platta ut ljusfältskanaler eller andra kanaler med varierande skärpedjup. Justera ljusstyrkan och kontrasten på bilderna med Bild | Justera | Ljusstyrka/kontrast. Använd skjutreglagen eller inmatningsfälten för att justera parametrarna och klicka sedan på Använd. Du kan också använda en sträckningsmetod för linjärt histogram genom att välja Process | Förbättra kontrasten …. Ange procentandelen mättade pixlar, välj Normalisera och klicka sedan på OK.OBS: När du kvantifierar eller jämför fluoroforsignalintensiteten mellan bilder är det i allmänhet bäst att inte justera ljusstyrkan eller kontrasten eftersom detta kan ändra de relativa pixelintensiteterna i bilden, vilket kan påverka mätningarna och jämförelserna. Om du vill skilja mellan kanaler använder du pseudofärger på specifika bilder med hjälp av en uppslagstabell (LUT). Det gör du genom att markera bilden och sedan välja Bild på menyn ImageJ/FIJI, klicka på Slå upp tabeller och välja lämplig LUT i listrutan. På menyn väljer du Bild | Färg | Slå samman kanaler. För varje kanal (C1, C2, C3…) använder du listrutan för att välja den bild som ska tilldelas den kanalen. Klicka på OK för att slå samman kanalerna. Spara den nya bilden som TIFF genom att välja Arkiv | Spara som | Tiff. Ange ett namn för filen och klicka på Spara.