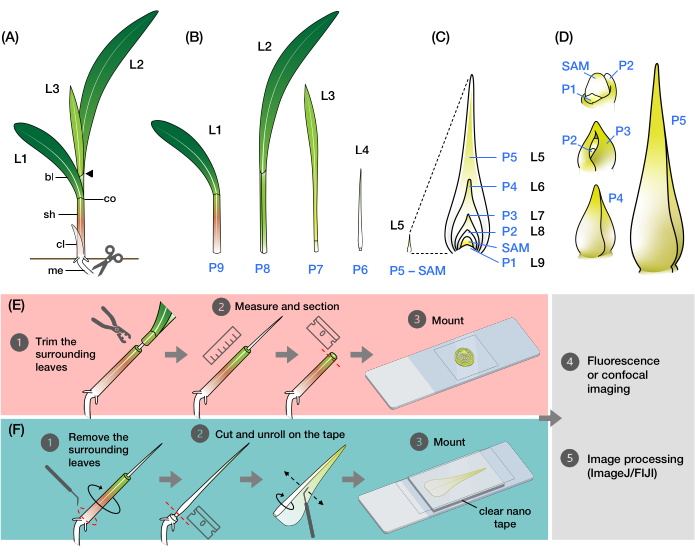
1. Iscenesættelse af majsbladudvikling og design af eksperimentet Dyrkning af planterne og iscenesættelse af bladudviklingBestem alder og udviklingsstadium for de planter, der skal bruges til eksperimentet, og udfør en detaljeret iscenesættelse af bladudvikling.BEMÆRK: Det anbefales, at iscenesættelsen udføres på planter med samme genetiske baggrund som de mutante eller transgene linjer, der vil blive brugt i forsøgene. SAM stopper med at producere nye blade mellem 20 og 25 DAG, afhængigt af den genetiske baggrund og vækstbetingelser. Af denne grund er metoderne beskrevet her ideelle til majsplanter i alderen 7-14 DAG. Dyrk majsplanterne i et drivhus eller vækstkammer efter en passende planteplejeprotokol35. Saml planterne i den ønskede alder eller V-trin med en lille kniv eller saks ved at skære ved mesocotyl, som er stammen under jordoverfladen (figur 1A). Indsæt forsigtigt skæreværktøjet i jorden ved siden af frøplanten og skub det ned til bunden i en vinkel på 45 ° for at skære mesocotyl. Løft frøplanten ud af jorden og støv eventuelt resterende snavs eller jordpartikler væk, der kan klamre sig til planten. Fjern coleoptilen, den beskyttende kappe, der dækker det nye skud i unge frøplanter, manuelt. For hver plante skal du registrere V-trinnet, antallet af blade og længden af det sidste blad, der kommer ud af hvirvlen. Tag billeder af planterne til fremtidig reference. Fjern de ældre blade en efter en. For at gøre dette skal du holde planten ved resterne af mesocotyl eller stamme og skære hvert blad fra bunden af kappen og forsigtigt folde kappen ud på en cirklende måde med en tandsonde (figur 1B).BEMÆRK: Se trin 2.1 for en hurtig metode til fjernelse af de omgivende blade. Under et stereomikroskop ved ca. 2x forstørrelse dissekeres resten af skudspidsen forsigtigt på fugtige servietter for at forhindre, at prøven tørrer ud. Skær forsigtigt og fold forsigtigt hvert bladprimordium ud med en tandsonde med en bøjet nål eller et par fine pincet, indtil SAM, P1 og P2 er synlige (figur 1D). Optag udseendet og målingen af hver plastochron.BEMÆRK: Se et eksempel på bladudviklingsstadietrin i majsplanter i supplerende fil 1. Planlægning af eksperimentetBEMÆRK: Planlægning af eksperimentet omhyggeligt kan forbedre effektiviteten. For eksempel i tværgående sektionsanalyse kan bestemmelse af den omtrentlige placering, der skal skæres, når der afbildes specifikke plastokroner eller regioner, reducere dissektionstiden. Figur 2 viser et eksempel på standardiseret prøveudtagning. Desuden kan optimering af billeddannelsesparametrene baseret på egenskaberne af FP’erne eller fluorescerende sonder forbedre effektiviteten af eksperimenterne.Brug iscenesættelsesoplysningerne som en guide til at fokusere eksperimentet på specifikke plastokroner, primordiumregioner og væv. Brug FP’erne eller andre fluorescerende sonder af interesse til at følge afsnit 2 og/eller 3 nedenfor på nogle få prøver.BEMÆRK: Se tilgængelige majs FP markør linje 36-42 frøbestande på https://www.maizegdb.org/data_center/stock. Bestem de optimale billedoptagelsesparametre for hver FP-reporter eller sonde, herunder den passende detektor eller filter, excitations- og emissionsbølgelængder, pinhole-størrelse og andre indstillinger. Gem indstillingerne for at gengive arbejdsvilkårene for hvert af eksperimenterne.BEMÆRK: Se Linh og Scarpella30 for nogle tekniske retningslinjer for konfokal laserscanningsmikroskopi af udviklende blade. Planlæg billedbehandlingstiden baseret på halveringstiden og andre egenskaber for FP reporter43.BEMÆRK: Tiden mellem dissektion og billeddannelse skal også overvejes ved planlægning af et forsøg, da friske planteprøver udsættes for udtørring og cellulære ændringer i denne periode. Figur 2: Prøveudtagningsplan til tværsnitsanalyse af majsbladprimordia. (A, venstre) Proksimalt skud af en 7 DAG majsplante, der viser et blotlagt fjerde blad (L4) ved P7. De brudte linjer angiver de syv prøveudtagningspunkter langs primordium, fra 0,5 mm til 10 mm. (A, højre) Skematisk gengivelse af bladets primordia med den projicerede størrelse og placering af hver plastochron: P7 (hvid); P6 (magenta); P5 (blå); P4 (grøn); og P3 indtil SAM (gul). (B-H) Fluorescensbilleder af tværsnit, der repræsenterer prøveudtagningsstederne vist i A fra 10 mm (B) til 0,5 mm (H). Primordierne er pseudofarvede i henhold til plastochronfarveskemaet i (A). Sektionerne blev afbildet med et epifluorescensmikroskop ved hjælp af et longpass-emission UV-filter til autofluorescens. Skalabjælke = 200 μm (B-H). Denne figur er blevet ændret og gengivet med tilladelse fra Robil og McSteen24. Forkortelser: DAG = dage efter spiring; P = plastochron; SAM = skyde apikal meristem; † = bladskede eller egentlig præ-ligul; * = skyde apikal meristem; ** = stilk. Klik her for at se en større version af denne figur. 2. Billeddannelse af tværsnit af majsbladprimordia Stripping af ældre omgivende blade og måling af bladets primordiumFrøplanten opsamles ved omhyggeligt at skære den ved mesocotylen under jordoverfladen med en lille kniv eller saks som beskrevet i trin 1.1.3-1.1.4 (figur 1A). Bestem den korrekte trådstripperhulstørrelse, der skal bruges, og hvor snittet skal foretages langs skuddet. Sørg for, at hullet er stort nok til at passe til målprimordium. Begynd at skære i den mere distale del af skuddet og arbejd gradvist mod basen, indtil spidsen af primordium er blottet. For at klippe skal du holde trådstripperen med kæberne vendt mod skuddet. Placer skuddet til det valgte hul, og klem stripperhåndtagene sammen for at skære gennem bladhvirvlen.BEMÆRK: For at undgå utilsigtet at skære målprimordium skal du forsigtigt justere midten af skuddet med hullet i trådstripperen. For større planter anbefales det manuelt at fjerne nogle af de omgivende blade manuelt, før du bruger stripperen.FORSIGTIG: Trådstrippere har skarpe kæber, der kan forårsage snit eller andre skader, hvis de ikke bruges forsigtigt. Mens du fortsætter med at klemme håndtagene sammen, skal du skubbe stripperen væk fra skuddet for at trimme den øverste del af bladene. Sørg for, at målområdet i primordium forbliver indhyllet af de omgivende blade (supplerende figur S1C, D).BEMÆRK: Efter trimning af de øverste blade efterlades skuddet med en udsat primordiumspids (P5, P6 eller P7, afhængigt af snittets position og størrelsen på det anvendte trådstripperhul; Figur 1E). Dette primordium vil tjene som reference for estimering af størrelsen og stadiet af yngre primordia. P6 er den mest praktiske reference til brug i majsplanter på grund af dets størrelsesområde og pinlignende morfologi. Brug en lineal til at måle længden fra spidsen af primordium til bunden af skuddet. Optag målingen.BEMÆRK: Primordiets længde vil blive lidt overvurderet, da der er et par millimeter stængel i bunden af primordien (figur 2A). Frihåndssnit og billeddannelse af bladprimordiumUnder et stereomikroskop med ca. 0,8x forstørrelse skal du holde optagelsen stabil på en glat overflade, såsom et glasskærebræt eller andet ridsefast materiale. Brug et barberblad eller skalpel til at skære tynde sektioner (0,25-0,8 mm) af skuddet på de ønskede punkter langs primordiumets længde. Lav et indledende snit lidt over målområdet for at kassere den distale del af skuddet, og få derefter tynde sektioner omkring målområdet. For at lave rene snit skal du holde bladet vinkelret på skuddet og skubbe det indad med en jævn, jævn bevægelse.BEMÆRK: Der kan være et par millimeter snitsrum for større primordia (P6 eller P7). For mindre primordier (P5 og tidligere) kan 1 mm dog gøre en betydelig forskel i prøveudtagningen, da disse primordia stables inden for de første par millimeter af skuddets bund (se figur 2A, F-H). Det anbefales at bruge et dobbeltkantet barberblad til finere sektionering.FORSIGTIG: Vær forsigtig, når du bruger barberblade og skalpeller for at undgå utilsigtede snit eller skader. Udskift barberbladet regelmæssigt, da bladskederne let kan sløve bladet. Dæk skuddet med fugtige klude efter hver sektionsrunde for at forhindre, at prøven tørrer ud. Monter bladsektionen på et rent glasglas (75 mm x 25 mm), brug en pipette til at påføre en dråbe vand (eller 50% glycerol) direkte på sektionen, og læg en dæksel (22 mm x 22 mm) ovenpå. Hvis du bruger et fluorescerende farvestof, skal du anvende farvestofopløsningen på sektionen, placere et dæksel ovenpå og inkubere det efter behov.BEMÆRK: Afhængigt af farvestoftypen kan yderligere trin til behandling af bladsektionerne være nødvendige, før du fortsætter til næste trin. Disse referencer44,45,46,47 giver generelle detaljer og anvendelser af fluorescerende farvestoffer i plantecellebilleddannelse. Et kernefarvningsfluorescerende farvestof, 5-ethynyl-2′-deoxyuridin (EdU), er blevet anvendt effektivt på tværsnit af majsbladprimordia48. I modsætning hertil giver plasmamembranfarvningsfarvestoffer, såsom Fei Mao 4-64 (FM 4-64) og propidiumiodid, ikke tilfredsstillende resultater (figur 3A-D). Placer diaset på scenen i et epifluorescens- eller konfokal laserscanningsmikroskop, og juster fokus og indstillinger efter behov for at visualisere fluoroforen30. Se tabel 1 for de belysnings- og billedoptagelsesindstillinger, der bruges til udvalgte FP-reporterlinjer for majs.BEMÆRK: Tykke sektioner kan producere høje niveauer af baggrundsautofluorescens. Overvej nogle billeddannelses- og prøveforberedelsesstrategier, der kan hjælpe med at afbøde dette problem30,49 (se tabel 2).FORSIGTIG: Vær forsigtig og brug beskyttelsesbriller, når du arbejder med kraftige lyskilder og lasere for at reducere risikoen for øjenskader. Billede gennem bladsektionen ved forskellige forstørrelser og detekteringskanaler (herunder lysstyrken)30. Brug 4x forstørrelse til at afbilde hele tværsnittet og 20x eller 40x forstørrelse til at afbilde specifikke væv. Reducer eksponeringstiden ved højere forstørrelser for epifluorescensbilleddannelse for at beskytte prøverne mod hurtig fotoblegning (se tabel 1). Tag billeder af hele sektionen eller bestemte interesseområder (ROI’er). Hvis det er nødvendigt, erhverve en z-stack, som er en række billeder taget på forskellige brændvidder30. For at erhverve en z-stak skal du først bestemme de øvre og nedre positioner i prøven og derefter antallet af optiske sektioner, der skal fanges mellem disse positioner. Færre optiske sektioner resulterer i en større z-trinstørrelse, som er afstanden mellem hver sektion. Afhængigt af FP’ens egenskaber og den anvendte forstørrelse optages tre til 25 optiske sektioner med z-trinstørrelser mellem 1 og 12 μm til billeddannelse af tværsnit af majsbladprimordia.BEMÆRK: For at samkopiere z-stakken skal du bruge en passende volumengengivelsesteknik50 , der kan udføres via mikroskopsoftwaren eller ved hjælp af ImageJ/FIJI34 (se trin 4.1 og 4.2). Gem alle billedfilerne og deres tilknyttede metadata, hvis de er tilgængelige. Tabel 1: Indstillinger for belysning og billedoptagelse, der anvendes til fluorescens og konfokal billeddannelse af udvalgte FP-rapportører for majs. Forkortelser: FP = fluorescerende protein; TRITC = tetramethylrhodamin FITC = fluoresceinisothiocyanat; WLL = laser med hvidt lys; Ar-ion = argonionlaser; HyD = hybrid detektor; AU = Luftig enhed; Hz = Hertz, scanningslinje pr. Sekund. Klik her for at downloade denne tabel. 3. Billeddannelse af valsede hele monteringer af majsbladprimordia Dissekering af primordia og klargøring af glasobjektglasset med nanotapeFrøplanten samles ved omhyggeligt at skære den ved mesocotylen under jordoverfladen med en lille kniv eller en saks som beskrevet i trin 1.1.3-1.1.4 (figur 1A). Forbered glasglasset (75 mm x 25 mm) med det klare, dobbeltsidede nanobånd (supplerende figur S1H), før du dissekerer planten. Skær et rektangulært stykke nanotape og sæt det fast i midten af et rent glasglas. Fjern ikke den beskyttende plastfilm på oversiden af båndet.BEMÆRK: Bestem størrelsen på båndet, der skal bruges, i henhold til størrelsen af prøverne til billedet (se eksempler på overdimensionerede prøver i supplerende figur S1I, J). Nanobåndet fås i enten akryl- eller polyurethangelmateriale. Begge typer er effektive til fluorescens og konfokal billeddannelse24. For at forhindre misfarvning af båndet skal du beskytte det mod langvarig udsættelse for lys ved at opbevare det i en mørk beholder. Begynd at dissekere skuddet ved at fjerne den øverste del af de ældre blade med en trådfjerner, som beskrevet i trin 2.1. Hold skuddet ved mesocotyl og fjern forsigtigt de omgivende blade med en tandsonde for at udtrække primordia. For at gøre dette skal du fjerne bladene fra bunden af skuddet og folde dem ud en ad gangen, indtil primordium af interesse er udsat. Alternativt kan du bruge trådstripperen til direkte at trække bladets primordia ud ved at lave et snit i skuddets basale område (supplerende figur S1E-G).BEMÆRK: Denne metode medfører en højere risiko for at snappe primordia. Montering og billeddannelse af primordium.Fjern beskyttelsesfilmen fra båndet. Læg det udsatte primordium på båndet. Brug et barberblad til at skære primordium i bunden, kassér basalstammen og hypocotyl (figur 1F). Rul primordium ud ved hjælp af en tandsonde med en bøjet nål (spidsdiameter på 0,25-0,6 mm). Brug en mikrosonde (spidsdiameter på 0,15-0,2 mm) til primordia mindre end 3 mm (P3 til tidlig P4). Placer nålespidsen parallelt med overfladen, fold den ydre basalmargen ud, og tryk den forsigtigt mod tapen. For en glattere udrulning smøres bladets indre overflade (adaxial) ved at dyppe nålens spids i 100% glycerol. Med den ydre margen klæbende til tapen justeres nålespidsen parallelt med bladets lange akse. Skub forsigtigt kanylen sidelæns for at rulle den ud, og flad bladet på tapen (figur 1F).BEMÆRK: Det er almindeligt at knække eller beskadige bladet under rullning, især i større primordia med en robust midrib (se figur 3E-G). Udrullning af bladet kraftigt kan blå mærke overfladen og producere artefakter under billeddannelse (figur 3H). Se tabel 2 og diskussionen for anbefalede løsninger på disse problemer. Påfør en dråbe vand på det urullede primordium. Placer straks en dæksel oven på vanddråben og primordium. Tryk forsigtigt kanterne af dækslet ned, så de klæber til båndet.BEMÆRK: Det anbefales at bruge store rektangulære dæksler (50 mm x 24 mm eller 60 mm x 24 mm) med ekstra områder til fastgørelse af tapen. Minimer dannelsen af luftbobler, som kan forvrænge bladet og forårsage ujævn lysbrydning (se figur 3I-K). Sørg for, at dæksedlen klæber helt til tapen, da kanterne på det monterede primordium kan rulle tilbage under et løst klæbet dæksel (se figur 3L). Placer objektglasset på scenen for et epifluorescens (se supplerende figur S1K) eller konfokal laserscanningsmikroskop, og juster fokus og indstillinger efter behov for at visualisere fluoroforen30. Se tabel 1 for de belysnings- og billedoptagelsesindstillinger, der bruges til de valgte rammerapportlinjer for majs. Afbild hele bladet eller et specifikt investeringsafkast gennem forskellige detektionskanaler30. For at afbilde hele bladet skal du enten manuelt tage mosaikbilleder af bladet eller bruge mikroskopets flisefunktion ved lav forstørrelse (4x eller 10x).BEMÆRK: Selv ved den laveste forstørrelse kan majsbladprimordia være for stor til at blive afbildet med et epifluorescens- eller konfokal laserscanningsmikroskop, hvilket resulterer i længere billedbehandlingstider, der kan resultere i fotoblegning (se figur 3M) eller øget baggrundsautofluorescens. Derfor anbefales det at bruge et fluorescensstereomikroskop til billeddannelse af helbladsprøver større end 5 mm. Til konfokal billeddannelse er det nødvendigt at opnå z-stakke, der omfatter bladets tykkelse (se trin 2.2.9). Gem alle billedfilerne og deres tilknyttede metadata, hvis de er tilgængelige. Tabel 2: Fejlfinding af almindelige problemer i billeddannelse af tværsnit og hele monteringer af majsbladprimordia. Klik her for at downloade denne tabel. Figur 3: Suboptimal tværsnit og helmonteret forberedelse af majsbladprimordia. (A-D) Repræsentative konfokale billeder af tværsnit af bladprimordia med en plasmamembranmarkør, PIP2-1-CFP (A), og et plasmamembranbindende fluorescerende farvestof, FM 4-64 (B), med tilsvarende lysfeltbilleder (C,D). Sammenlignet med PIP2-1-CFP viser FM 4-64 suboptimal visualisering af cellekonturer. (E-M) Repræsentative fluorescensbilleder af hele monteringer af bladprimordier, der viser tilstedeværelsen af rivning (E-G), forslåede overflader† (H), luftbobler* (I-K), tilbagerullede margener (L) og fotoblegede områder‡ (M). Bladets primordia udtrykker DII-Venus (E-G), GAR2-YFP (H-J), mDII-Venus (K), PIN1a-YFP (L) og DR5-RFP (M). Skalabjælke = 200 μm (A-D); 500 μm (E-M). Figur 3A er blevet ændret og gengivet med tilladelse fra Robil og McSteen24, mens figur 5B-M er upublicerede data fra forfatterne. Forkortelser: P = plastochron; YFP = gult fluorescerende protein; RFP = rødt fluorescerende protein; CFP = cyanfluorescerende protein PIP2-1-CFP = p Zm PIP2-1::ZmPIP2-1:CFP; DII-Venus = pZmUbi:DII:YFP-NLS; GAR2-YFP = p Zm GAR2::ZmGAR2:YFP; mDII-Venus = pZmUbi:mDII:YFP-NLS; PIN1a-YFP = p Zm PIN1a::ZmPIN1a:YFP; DR5-RFP = DR5rev::mRFPer; BF = lysfelt. Klik her for at se en større version af denne figur. 4. Behandling af billederne ved hjælp af ImageJ / FIJI Samkopiering af z-stakke ved hjælp af maksimal intensitetsprojektion (MIP)Start ImageJ/FIJI , og åbn stakken med billeder, eller opret en ny stak ud fra flere separate billeder ved at vælge Billede | Stakke | Billeder, der skal stables i menuen. Vælg billedstakken. Vælg Billede | i menuen Stakke | Z-projekt…. I dialogboksen Z-projekt skal du vælge Maks. intensitet i rullemenuen Projektionstype . Klik på OK for at oprette z-projektionen med maksimal intensitet.BEMÆRK: Det samkopierede billede er en 2D-projektion af pixel med den højeste intensitet på tværs af z-stakkene50. Denne metode er egnet til visualisering af fluoroforen, men ikke de anatomiske træk i lyse feltbilleder af tykke tværgående sektioner og helbladsmonteringer. Gem billedet som TIFF (Tagged Image File Format) ved at vælge Filer | Gem som | Tiff. Indtast et navn til filen, og klik på Gem.BEMÆRK: Det anbefales at bruge TIFF til lagring af fluorescens og konfokale billeder, da det bevarer kvaliteten og detaljerne, når billedet gemmes. Samkopiering af z-stakke ved hjælp af EDF-plugin (Extended Depth of Field)51Installer EDF-plugin52. Download Extended_Depth_Field.jar filen (version 17.05.2021) fra http://bigwww.epfl.ch/demo/edf/, og placer den i mappen “plugins” i ImageJ/FIJI. Start ImageJ/FIJI , og åbn stakken med billeder, eller opret en ny stak ud fra flere separate billeder ved at vælge Billede | Stakke | Billeder, der skal stables i menuen. Vælg billedstakken. I menuen skal du vælge Plugins | Udvidet dybdeskarphed | EDF nem tilstand. I dialogboksen Udvidet dybdeskarphed skal du bruge skyderne under Hastighed/ kvalitet til at indstille den ønskede kvalitet og behandlingshastighed og Højdekortreg til at indstille udjævningsniveauet for topografien. Klik på Kør for at oprette det rekonstruerede billede.BEMÆRK: EDF kombinerer stakken til et skarpt, sammensat 2D-billede ved at projicere det bedste fokus51. Denne teknik er nyttig til at overvinde den begrænsede dybdeskarphed og er velegnet til rekonstruktion af lysfeltsbilleder af tykke tværgående sektioner eller helbladsmonteringer. Vær forsigtig, når du anvender dette filter til at visualisere fluoroforen, fordi EDF i modsætning til MIP justerer billedets kontrast og påvirker pixelintensiteten. Gem billedet som en TIFF-fil ved at vælge Filer | Gem som | Tiff. Indtast et navn til filen, og klik på Gem. Syning af billeder sammen ved hjælp af Grid / Collection Stitching-plugin53Gem samlingen af billeder eller billedstakke, der skal sys sammen i en enkelt mappe. Start ImageJ / FIJI, og vælg Plugins | Syning | Gitter-/samlingssyning fra menuen. I dialogboksen Gitter-/samlingssømning skal du vælge Ukendt position i rullemenuen Type og derefter Alle filer i mappen fra Rækkefølge. Klik på OK. I dialogboksen Gittersyning: Ukendt placering, Alle filer i mappen skal du angive placeringen af billederne ved at skrive eller indsætte mappestien i tekstfeltet Mappe eller ved at klikke på Gennemse… for at finde mappen. Klik på OK for at starte syningsprocessen.BEMÆRK: Indstillingen Ukendt position i plugin’et Grid/Collection Stitching rekonstruerer et sammensat billede ud fra et sæt sammenhængende billeder ved at analysere deres overlappende områder. Derfor er denne mulighed nyttig til syning af mosaikker af billeder af tværgående sektioner eller helbladsmonteringer, der ikke opnås ved hjælp af en flisescanning. Når syningen er færdig, vises et nyt vindue med det syede billede. Gem billedet som en TIFF-fil ved at vælge Filer | Gem som | Tiff. Indtast et navn til filen, og klik på Gem.BEMÆRK: Syning af store montager af 2D- og 3D-billeder kan også udføres ved hjælp af billedrekonstruktionskommandoer i mikroskopsoftwaren eller via Bio-Formats Import Options i ImageJ / FIJI. Kombinere flere kanaler ved hjælp af kommandoen Flet kanalerStart ImageJ / FIJI, og åbn de billeder eller billedstakke, der skal flettes.BEMÆRK: Billedstakke kan først samkopieres ved hjælp af trin 4.1 eller 4.2. MIP anbefales generelt til udfladning af fluoroforkanaler, mens EDF er bedre egnet til udfladning af lysfeltkanaler eller andre kanaler med varierende fokusdybde. Juster billedernes lysstyrke og kontrast med Image | Juster | Lysstyrke/kontrast. Brug skyderne eller indtastningsfelterne til at justere parametrene, og klik derefter på Anvend. Du kan også bruge en lineær histogramstrækningsmetode ved at vælge Proces | Forbedr kontrasten…. Indstil procentdelen af mættede pixel, vælg Normaliser, og klik derefter på OK.BEMÆRK: Når du kvantificerer eller sammenligner fluoroforsignalintensiteten mellem billeder, er det generelt bedst ikke at justere lysstyrken eller kontrasten, da dette kan ændre de relative pixelintensiteter i billedet, hvilket kan påvirke målingerne og sammenligningerne. Hvis du vil skelne mellem kanaler, skal du anvende pseudofarver på bestemte billeder ved hjælp af en opslagstabel (LUT). For at gøre dette skal du vælge billedet og derefter vælge Billede i menuen ImageJ/FIJI, klikke på Opslagstabeller og vælge den relevante LUT på rullelisten. Vælg Billede | i menuen Farve | Flet kanaler. For hver kanal (C1, C2, C3…) skal du bruge rullelisten til at vælge det billede, der skal tildeles den pågældende kanal. Klik på OK for at flette kanalerne. Gem det nye billede som TIFF ved at vælge Filer | Gem som | Tiff. Indtast et navn til filen, og klik på Gem.