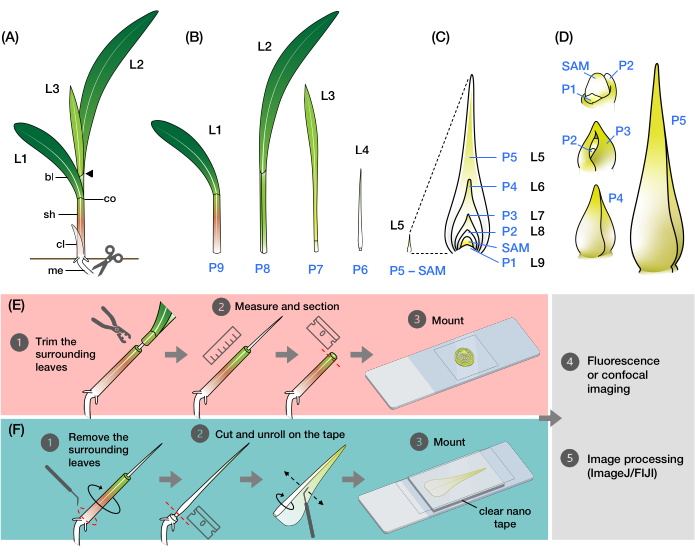
1. Iscenesettelse av maisbladutvikling og utforming av eksperimentet Dyrking av plantene og iscenesettelse av bladutviklingBestem alder og utviklingsstadium av plantene som skal brukes til forsøket, og utfør en detaljert iscenesettelse av bladutvikling.MERK: Det anbefales at staging utføres på planter med samme genetiske bakgrunn som de mutante eller transgene linjene som skal brukes i forsøkene. SAM slutter å produsere nye blader mellom 20 og 25 DAG, avhengig av genetisk bakgrunn og vekstforhold. Av denne grunn er metodene beskrevet her ideelle for maisplanter i alderen 7-14 DAG. Dyrk maisplantene i et drivhus eller vekstkammer etter en egnet plantepleieprotokoll35. Samle plantene i ønsket alder eller V-stadium med en liten kniv eller saks ved å kutte på mesocotyl, som er stammen under jordoverflaten (figur 1A). Sett skjæreverktøyet forsiktig i jorden ved siden av frøplanten og skyv det ned til basen i en 45 ° vinkel for å kutte mesokotylen. Løft frøplanten ut av jorden og støv bort gjenværende smuss eller jordpartikler som kan klamre seg til planten. Fjern coleoptile, beskyttelseskjeden som dekker det fremvoksende skuddet i unge frøplanter, for hånd. For hver plante registrerer du V-trinnet, antall blader og lengden på det siste bladet som kommer fra hvirvelen. Ta bilder av plantene for fremtidig referanse. Fjern de eldre bladene en etter en. For å gjøre dette, hold planten ved restene av mesocotyl eller stammen, og punkter hvert blad fra bunnen av skjeden og forsiktig unfurl skjeden på en sirklende måte med en tannsonde (figur 1B).MERK: Se trinn 2.1 for en rask metode for å fjerne de omkringliggende bladene. Under et stereomikroskop ved rundt 2x forstørrelse, dissekerer du forsiktig resten av skuddet på fuktige kluter for å forhindre at prøven tørker ut. Stikk forsiktig ut og brett ut hvert blad primordium med en tannsonde med en bøyd nål eller et par fine tang til SAM, P1 og P2 er synlige (figur 1D). Registrer utseendet og måling av hver plastokron.MERK: Se et eksempel på bladutviklingsstaging hos maisplanter i tilleggsfil 1. Planlegging av eksperimentetMERK: Å planlegge eksperimentet nøye kan forbedre effektiviteten. For eksempel, i tverrsnittanalyse, kan bestemmelse av omtrentlig plassering som skal kuttes ved avbildning av spesifikke plastokroner eller regioner, redusere disseksjonstiden. Figur 2 viser et eksempel på standardisert prøvetaking. Videre kan optimalisering av avbildningsparametrene basert på egenskapene til FPs eller fluorescerende sonder forbedre effektiviteten av eksperimentene.Bruk iscenesettelsesinformasjonen som en veiledning for å fokusere eksperimentet på spesifikke plastokroner, primordiumregioner og vev. Bruk FPs eller andre fluorescerende sonder av interesse for å følge avsnitt 2 og / eller 3 nedenfor på noen få prøver.MERK: Se tilgjengelige mais FP markør line36-42 frø aksjer på https://www.maizegdb.org/data_center/stock. Bestem de optimale parameterne for bildeopptak for hver FP-reporter eller sonde, inkludert passende detektor eller filter, eksitasjons- og utslippsbølgelengder, pinhole-størrelse og andre innstillinger. Lagre innstillingene for å gjengi arbeidsforholdene for hvert av eksperimentene.MERK: Se Linh og Scarpella30 for noen tekniske retningslinjer for konfokal laserskanningsmikroskopi av utviklende blader. Planlegg bildetiden basert på halveringstiden og andre egenskaper til FP reporter43.MERK: Tiden mellom disseksjon og avbildning må også vurderes når du planlegger et eksperiment, da ferske planteprøver blir utsatt for uttørking og cellulære endringer i denne perioden. Figur 2: Prøvetakingsskjema for tverrsnittsanalyse av maisbladprimordia. (A, venstre) Proksimalt skudd av en 7 DAG maisplante, som viser et eksponert fjerde blad (L4) på P7. De brutte linjene indikerer de syv prøvetakingspunktene langs primordium, fra 0, 5 mm til 10 mm. (A, høyre) Skjematisk fremstilling av bladprimordia, med den projiserte størrelsen og posisjonen til hver plastokron: P7 (hvit); P6 (magenta); P5 (blå); P4 (grønn); og P3 til SAM (gul). (VG Nett) Fluorescensbilder av tverrsnitt som representerer prøvetakingspunktene vist i A fra 10 mm (B) til 0,5 mm (H). Primordiene er pseudofarget i henhold til plastokronfargeskjemaet i (A). Snittene ble avbildet med et epifluorescensmikroskop ved hjelp av et UV-filter med longpass-emisjon for autofluorescens. Skala bar = 200 μm (B-H). Denne figuren er modifisert og gjengitt med tillatelse fra Robil og McSteen24. Forkortelser: DAG = dager etter spiring; P = plastokron; SAM = skyte apikal meristem; † = bladkappe eller pre-ligule riktig; * = skyte apikal meristem; ** = stamme. Klikk her for å se en større versjon av denne figuren. 2. Avbildning av tverrsnitt av maisblad primordia Stripping eldre omkringliggende blader og måling av bladet primordiumSamle frøplanten ved å kutte den forsiktig ved mesokotylen under jordoverflaten med en liten kniv eller saks, som beskrevet i trinn 1.1.3-1.1.4 (figur 1A). Bestem riktig wire stripper hullstørrelse som skal brukes og hvor kuttet skal gjøres langs skytingen. Forsikre deg om at hullet er stort nok til å passe til målprimordiumet. Begynn å kutte på den mer distale delen av skuddet og arbeid gradvis mot basen til spissen av primordiet er eksponert. For å gjøre kuttet, hold trådstripperen med kjevene mot skuddet. Plasser skuddet til det valgte hullet og klem stripperhåndtakene sammen for å skjære gjennom bladhvirvelen.MERK: For å unngå å kutte målprimordiet ved et uhell, juster midten av skuddet forsiktig med hullet i trådstripperen. For større planter anbefales det å manuelt fjerne noen av de omkringliggende bladene for hånd før du bruker stripperen.FORSIKTIG: Wire strippers har skarpe kjever som kan forårsake kutt eller andre skader hvis de ikke brukes forsiktig. Mens du fortsetter å klemme håndtakene sammen, skyver du stripperen bort fra skuddet for å trimme den øvre delen av bladene. Sørg for at målområdet i primordium forblir omsluttet av de omkringliggende bladene (tilleggsfigur S1C,D).MERK: Etter å ha trimmet de øvre bladene, sitter skuddet igjen med en eksponert primordiumspiss (P5, P6 eller P7, avhengig av kuttets posisjon og størrelsen på trådstripperhullet som brukes; Figur 1E). Dette primordium vil tjene som referanse for å estimere størrelsen og stadiet av yngre primordia. P6 er den mest praktiske referansen å bruke i maisplanter på grunn av størrelsesområdet og pin-lignende morfologi. Bruk en linjal til å måle lengden fra spissen av primordium til bunnen av skuddet. Registrer målingen.MERK: Lengden på primordium vil bli litt overestimert da det er noen få millimeter stamme ved foten av primordia (figur 2A). Frihåndssnitt og avbildning av bladprimordiumUnder et stereomikroskop med rundt 0,8x forstørrelse, hold skuddet stødig på en jevn overflate, for eksempel et skjærebrett i glass eller annet ripebestandig materiale. Bruk et barberblad eller skalpell til å kutte tynne seksjoner (0,25-0,8 mm) av skuddet på de ønskede punktene langs primordiumets lengde. Gjør et første kutt litt over målområdet for å forkaste den distale delen av skytingen, og få deretter tynne seksjoner rundt målområdet. For å gjøre rene kutt, hold bladet vinkelrett på skuddet og skyv det innover med en jevn, jevn bevegelse.MERK: Det kan være noen millimeter seksjoneringsrom for større primordia (P6 eller P7). For mindre primordia (P5 og tidligere) kan imidlertid 1 mm utgjøre en betydelig forskjell i prøvetakingen, da disse primordiene er stablet opp innenfor de første millimeterne av skuddets base (se figur 2A, F-H). Det anbefales å bruke et dobbeltkantet barberblad for finere seksjonering.FORSIKTIG: Vær forsiktig når du bruker barberblader og skalpeller for å unngå utilsiktede kutt eller skader. Bytt ut barberbladet regelmessig, siden bladkappene lett kan sløve bladet. Dekk skuddet med fuktige kluter etter hver seksjonsrunde for å forhindre at prøven tørker ut. Monter bladseksjonen på en ren glassglass (75 mm x 25 mm), bruk en pipette til å påføre en dråpe vann (eller 50% glyserol) direkte på seksjonen, og legg en deksel (22 mm x 22 mm) på toppen. Hvis du bruker et fluorescerende fargestoff, må du bruke fargestoffløsningen på seksjonen, plassere en dekselslip på toppen og inkubere den etter behov.MERK: Avhengig av fargestofftypen kan det være nødvendig med ytterligere trinn for behandling av bladseksjonene før du går videre til neste trinn. Disse referansene44,45,46,47 gir generelle detaljer og bruk av fluorescerende fargestoffer i plantecelleavbildning. Et kjernefargende fluorescerende fargestoff, 5-etynyl-2′-deoksyuridin (EdU), har blitt brukt effektivt på tverrsnitt av maisbladprimordia 48. Derimot gir plasmamembranfargestoffer, som Fei Mao 4-64 (FM 4-64) og propidiumjodid, ikke tilfredsstillende resultater (figur 3A-D). Plasser lysbildet på scenen av et epifluorescens eller konfokal laserskanningsmikroskop og juster fokus og innstillinger etter behov for å visualisere fluoroforen30. Se tabell 1 for innstillinger for belysning og bildeopptak som brukes for utvalgte mais FP reporter linjer.MERK: Tykke seksjoner kan produsere høye nivåer av bakgrunnsautofluorescens. Vurder noen bildebehandlings- og prøveprepareringsstrategier som kan bidra til å redusere dette problemet 30,49 (se tabell 2).FORSIKTIG: Vær forsiktig og bruk vernebriller når du arbeider med kraftige lyskilder og lasere for å redusere risikoen for øyeskader. Bilde gjennom bladsnittet ved forskjellige forstørrelser og deteksjonskanaler (inkludert belysningen i det lyse feltet)30. Bruk 4x forstørrelse til å avbilde hele tverrsnittet og 20x eller 40x forstørrelse for å avbilde spesifikke vev. Reduser eksponeringstiden ved høyere forstørrelser for epifluorescensavbildning for å beskytte prøvene mot rask fotobleking (se tabell 1). Ta bilder av hele delen eller bestemte interesseområder (ROI). Skaff deg om nødvendig en z-stack, som er en serie bilder tatt med forskjellige brennvidder30. For å skaffe deg en z-stack, bestem først de øvre og nedre posisjonene i prøven, deretter antall optiske seksjoner som må fanges mellom disse posisjonene. Færre optiske seksjoner resulterer i en større z-trinnstørrelse, som er avstanden mellom hver seksjon. Avhengig av egenskapene til FP og forstørrelsen som brukes, fang tre til 25 optiske seksjoner med z-trinnstørrelser mellom 1 og 12 μm for avbildning av tverrsnitt av maisbladprimordia.MERK: Hvis du vil flate ut z-stakken, bruker du en passende volumgjengivelsesteknikk50 som kan oppnås gjennom mikroskopprogramvaren eller ved å bruke ImageJ/FIJI34 (se trinn 4.1 og 4.2). Lagre alle bildefilene og de tilknyttede metadataene, hvis tilgjengelig. Tabell 1: Innstillinger for belysning og bildeopptak brukt til fluorescens og konfokal avbildning av utvalgte mais-FP-reportere. Forkortelser: FP = fluorescerende protein; TRITC = tetrametylrhodamin; FITC = fluoresceinisotiocyanat; WLL = hvitt lys laser; Ar-ion = argonionlaser; HyD = hybrid detektor; AU = Luftig enhet; Hz = Hertz, skannelinje per sekund. Klikk her for å laste ned denne tabellen. 3. Avbildning av utrullede hele fester av maisblad primordia Dissekere primordia og forberede glassglasset med nanotapeSamle frøplanten ved å kutte den forsiktig på mesocotyl under jordoverflaten med en liten kniv eller en saks, som beskrevet i trinn 1.1.3-1.1.4 (figur 1A). Forbered glasssliden (75 mm x 25 mm) med den klare, dobbeltsidige nanotapen (tilleggsfigur S1H) før du dissekerer planten. Klipp et rektangulært stykke nanotape og fest det til midten av et rent glassglass. Ikke fjern den beskyttende plastfilmen på oversiden av båndet.MERK: Bestem størrelsen på båndet som skal brukes i henhold til størrelsen på prøvene til bildet (se eksempler på store prøver i tilleggsfigur S1I, J). Nanobåndet er tilgjengelig i enten akryl eller polyuretan gel materiale. Begge typer er effektive for fluorescens og konfokal avbildning24. For å forhindre misfarging av båndet, beskytt det mot langvarig eksponering for lys ved å lagre det i en mørk beholder. Begynn å dissekere skuddet ved å fjerne den øvre delen av de eldre bladene med en trådstripper, som beskrevet i trinn 2.1. Hold skuddet ved mesocotyl og fjern forsiktig de omkringliggende bladene med en tannsonde for å trekke ut primordia. For å gjøre dette, fjern bladene fra bunnen av skuddet og utfold dem en om gangen til primordium av interesse er utsatt. Alternativt kan du bruke trådstripperen til å trekke ut bladprimordiene direkte ved å lage et kutt i basalområdet av skuddet (tilleggsfigur S1E-G).MERK: Denne metoden har en høyere risiko for å snappe primordia. Montering og avbildning av primordium.Fjern beskyttelsesfilmen fra båndet. Legg det eksponerte primordium på båndet. Bruk et barberblad til å skjære primordium ved basen, og kast basalstammen og hypocotyl (figur 1F). Rull ut primordium ved hjelp av en tannsonde med en bøyd nål (spissdiameter på 0,25-0,6 mm). Bruk en mikrosonde (spissdiameter på 0,15-0,2 mm) for primordia mindre enn 3 mm (P3 til tidlig P4). Plasser nålespissen parallelt med overflaten og brett ut den ytre basalmargen og press den forsiktig mot tapen. For jevnere utrulling, smør den indre overflaten (adaxial) av bladet ved å dyppe spissen av nålen i 100% glyserol. Med den ytre margen festet til båndet, juster nålespissen parallelt med bladets lange akse. Skyv kanylen forsiktig sidelengs for å rulle ut og flate bladet på tapen (figur 1F).MERK: Å bryte eller skade bladet mens du ruller ut er vanlig, spesielt i større primordia med en robust midrib (se figur 3E-G). Hvis du ruller bladet kraftig ut, kan det få blåmerker på overflaten og produsere artefakter under avbildning (figur 3H). Se tabell 2 og diskusjonen for anbefalte løsninger på disse problemene. Påfør en dråpe vann på det utrullede primordium. Legg umiddelbart en dekkslipp på toppen av vanndråpen og primordiumet. Trykk forsiktig ned kantene på dekselet slik at de fester seg til båndet.MERK: Det anbefales å bruke store rektangulære deksler (50 mm x 24 mm eller 60 mm x 24 mm) med ekstra områder for å feste båndet. Minimer dannelsen av luftbobler, som kan forvrenge bladet og forårsake ujevn lysbrytning (se figur 3I-K). Forsikre deg om at dekselet fester seg helt til båndet, da kantene på det monterte primordiumet kan rulle inn igjen under en løst festet dekselslip (se figur 3L). Plasser lysbildet på scenen av en epifluorescens (se tilleggsfigur S1K) eller konfokal laserskanningsmikroskop og juster fokus og innstillinger etter behov for å visualisere fluoroforen30. Se tabell 1 for innstillingene for belysning og bildeopptak som brukes for de valgte maisrapportlinjene. Avbilde hele bladet eller en bestemt avkastning gjennom ulike deteksjonskanaler30. For å avbilde hele bladet, må du enten manuelt ta mosaikkbilder av bladet eller bruke flisleggingen av mikroskopet ved lav forstørrelse (4x eller 10x).MERK: Selv ved den laveste forstørrelsen kan maisbladprimordia være for stor til å avbildes med et epifluorescens eller konfokal laserskanningsmikroskop, noe som resulterer i lengre bildebehandlingstider som kan føre til fotobleking (se figur 3M) eller økt bakgrunnsautofluorescens. Derfor anbefales det å bruke et fluorescensstereomikroskop for avbildning av helbladprøver større enn 5 mm. For konfokal avbildning vil det være nødvendig å oppnå z-stabler som omfatter tykkelsen på bladet (se trinn 2.2.9). Lagre alle bildefilene og de tilknyttede metadataene, hvis tilgjengelig. Tabell 2: Feilsøking av vanlige problemer ved avbildning av tverrsnitt og hele fester av maisbladprimordia. Klikk her for å laste ned denne tabellen. Figur 3: Suboptimalt tverrsnitt og helmontert preparering av maisbladprimordia. (A-D) Representative konfokale bilder av tverrsnitt av bladprimordia med en plasmamembranmarkør, PIP2-1-CFP (A), og et plasmamembranbindende fluorescerende fargestoff, FM 4-64 (B), med tilsvarende lysfeltbilder (C,D). Sammenlignet med PIP2-1-CFP, viser FM 4-64 suboptimal visualisering av cellekonturer. (E-M) Representative fluorescensbilder av hele fester av bladprimordier som viser tilstedeværelse av rifter (E-G), blåste overflater† (H), luftbobler* (I-K), tilbakerullede marger (L) og fotoblekede regioner‡ (M). Bladprimordia uttrykker DII-Venus (E-G), GAR2-YFP (H-J), mDII-Venus (K), PIN1a-YFP (L) og DR5-RFP (M). Skala bar = 200 μm (A-D); 500 μm (E-M). Figur 3A er modifisert og gjengitt med tillatelse fra Robil og McSteen24, mens figur 5B-M er upubliserte data fra forfatterne. Forkortelser: P = plastokron; YFP = gult fluorescerende protein; RFP = rødt fluorescerende protein; CFP = cyan fluorescerende protein; PIP2-1-CFP = pZmPIP2-1::ZmPIP2-1:CFP; DII-Venus = pzmubi:DII:YFP-NLS; GAR2-YFP = p Zm GAR2::ZmGAR2: YFP; mDII-Venus = pZmUbi:mDII:YFP-NLS; PIN1a-YFP = p Zm PIN1a::ZmPIN1a: YFP; DR5-RFP = DR5rev::mRFPer; BF = lyst felt. Klikk her for å se en større versjon av denne figuren. 4. Behandle bildene ved hjelp av ImageJ / FIJI Slå sammen z-stakker ved hjelp av maksimal intensitetsprojeksjon (MIP)Start ImageJ/FIJI og åpne bunken med bilder, eller opprett en ny stabel fra flere separate bilder ved å velge Image | Stabler | Bilder som skal stables fra menyen. Velg bildestakken. Fra menyen velger du Bilde | Stabler | Z-prosjekt …. I dialogboksen Z-Project velger du Maksimal intensitet fra rullegardinmenyen Projeksjonstype . Klikk OK for å opprette z-projeksjon med maksimal intensitet.MERK: Det sammenslåtte bildet er en 2D-projeksjon av pikslene med høyest intensitet over z-stakkene50. Denne metoden er egnet for å visualisere fluoroforen, men ikke de anatomiske egenskapene i lyse feltbilder av tykke tverrsnitt og helbladfester. Lagre bildet som TIFF (Tagged Image File Format) ved å velge File | Lagre som | Tiff. Skriv inn et navn på filen, og klikk Lagre.MERK: Det anbefales å bruke TIFF for lagring av fluorescens og konfokale bilder, da det beholder kvaliteten og detaljene når bildet lagres. Slå sammen z-stabler ved hjelp av Extended Depth of Field (EDF) plugin51Installer EDF-plugin52. Last ned Extended_Depth_Field.jar filen (versjon 17.05.2021) fra http://bigwww.epfl.ch/demo/edf/ og legg den i “plugins”-mappen til ImageJ/FIJI. Start ImageJ/FIJI og åpne bunken med bilder, eller opprett en ny stabel fra flere separate bilder ved å velge Image | Stabler | Bilder som skal stables fra menyen. Velg bildestakken. Fra menyen velger du Plugins | Utvidet dybdeskarphet | EDF enkel modus. I dialogboksen Utvidet dybdeskarphet bruker du glidebryterne under Hastighet/ kvalitet til å angi ønsket kvalitet og behandlingshastighet, og høydekartregeringen til å angi jevnhetsnivået for topografien. Klikk Kjør for å opprette det rekonstruerte bildet.MERK: EDF kombinerer stakken til et skarpt, sammensatt 2D-bilde ved å projisere det beste fokuset51. Denne teknikken er nyttig for å overvinne den begrensede fokusdybden og er egnet for å rekonstruere bilder av lyse felt av tykke tverrsnitt eller helbladfester. Vær forsiktig når du bruker dette filteret til å visualisere fluoroforen fordi EDF, i motsetning til MIP, justerer kontrasten i bildet, noe som påvirker pikselintensiteten. Lagre bildet som en TIFF-fil ved å velge Fil | Lagre som | Tiff. Skriv inn et navn på filen, og klikk Lagre. Sy bilder sammen ved hjelp av Grid / Collection Stitching plugin53Lagre samlingen av bilder eller bildestakker som skal sys sammen i en enkelt katalog. Start ImageJ/FIJI og velg Plugins | Søm | Rutenett/samlingssøm fra menyen. I dialogboksen Sammensetting av rutenett / samling velger du Ukjent plassering fra rullegardinmenyen Type og deretter Alle filer i katalogen fra Rekkefølge. Klikk OK. I dialogboksen Rutenettsammensetting: Ukjent plassering, Alle filer i katalog , angir du plasseringen av bildene ved å skrive eller lime inn katalogbanen i Katalogtekst-feltet eller ved å klikke Bla gjennom… for å finne katalogen. Klikk OK for å starte sammensyingsprosessen.MERK: Alternativet Ukjent posisjon i plugin-modulen Grid/Collection Stitching rekonstruerer et sammensatt bilde fra et sett med sammenhengende bilder ved å analysere overlappende regioner. Derfor er dette alternativet nyttig for å sy mosaikker av bilder av tverrsnitt eller helbladfester som ikke oppnås ved hjelp av en flisskanningsoperasjon. Når sømmen er ferdig, vises et nytt vindu med det sydde bildet. Lagre bildet som en TIFF-fil ved å velge Fil | Lagre som | Tiff. Skriv inn et navn på filen, og klikk Lagre.MERK: Sammensetting av store montasjer av 2D- og 3D-bilder kan også oppnås ved hjelp av bilderekonstruksjonskommandoer i mikroskopprogramvaren eller gjennom Bio-Formats Import Options i ImageJ / FIJI. Kombinere flere kanaler ved hjelp av kommandoen Slå sammen kanalerStart ImageJ/FIJI og åpne bildene eller bildestakkene som skal slås sammen.MERK: Bildestakker kan slås sammen først ved hjelp av trinn 4.1 eller 4.2. MIP anbefales generelt for å flate ut fluoroforkanaler, mens EDF er bedre egnet for å flate ut lysfeltkanaler eller andre kanaler med varierende dybdefokus. Juster lysstyrken og kontrasten til bildene med Image | Justere | Lysstyrke/kontrast. Bruk glidebryterne eller inndatafeltene til å justere parameterne, og klikk deretter på Bruk. Du kan også bruke en lineær histogramstrekkemetode ved å velge Prosess | Forbedre kontrasten …. Angi prosentandelen av mettede piksler, velg Normaliser, og klikk deretter OK.MERK: Når du kvantifiserer eller sammenligner fluoroforsignalintensiteten mellom bilder, er det vanligvis best å ikke justere lysstyrken eller kontrasten, fordi dette kan endre den relative pikselintensiteten i bildet, noe som kan påvirke målingene og sammenligningene. Hvis du vil skille mellom kanaler, bruker du pseudofarger på bestemte bilder ved hjelp av en oppslagstabell (LUT). For å gjøre dette, velg bildet, velg deretter Bilde fra ImageJ / FIJI-menyen, klikk Slå opp tabeller og velg riktig LUT fra rullegardinlisten. Fra menyen velger du Bilde | Farge | Slå sammen kanaler. For hver kanal (C1, C2, C3 …) bruker du rullegardinlisten til å velge bildet som skal tilordnes den kanalen. Klikk OK for å slå sammen kanalene. Lagre det nye bildet som TIFF ved å velge Fil | Lagre som | Tiff. Skriv inn et navn på filen, og klikk Lagre.