הדמיה של mtHyPer7, ביוסנסור רציומטרי למי חמצן מיטוכונדריאליים, בתאי שמרים חיים
Summary
מי חמצן (H 2 O2) הםגם מקור לנזקי חמצון וגם מולקולת איתות. פרוטוקול זה מתאר כיצד למדוד H 2 O2מיטוכונדריאלי באמצעות HyPer7 ממוקד מיטוכונדריה (mtHyPer7), ביוסנסור יחסי מקודד גנטית, בשמרים חיים. הוא מפרט כיצד לייעל את תנאי ההדמיה ולבצע אנליזה כמותית תאית ותת-תאית באמצעות תוכנה זמינה באופן חופשי.
Abstract
תפקוד לקוי של המיטוכונדריה, או שינוי תפקודי, נמצא במחלות ומצבים רבים, כולל מחלות נוירודגנרטיביות והפרעות שרירים ושלד, סרטן והזדקנות נורמלית. כאן מתוארת גישה להערכת תפקוד מיטוכונדריאלי בתאי שמרים חיים ברזולוציות תאיות ותת-תאיות באמצעות ביו-חיישן יחסי, זעיר פולשני ומקודד גנטית. הביוסנסור, HyPer7 ממוקד מיטוכונדריה (mtHyPer7), מזהה מי חמצן (H 2 O2) במיטוכונדריה. הוא מורכב מרצף אותות מיטוכונדריאלי המאוחה לחלבון פלואורסצנטי בעל מוטציה מעגלית ותחום H 2 O2המגיב לחלבון OxyR חיידקי. הביוסנסור נוצר ומשולב בגנום השמרים באמצעות מערכת נטולת סמנים CRISPR-Cas9, לביטוי עקבי יותר בהשוואה למבנים הנישאים בפלסמיד.
mtHyPer7 ממוקד כמותית למיטוכונדריה, אין לו השפעה ניתנת לזיהוי על קצב גדילת שמרים או מורפולוגיה מיטוכונדריאלית, ומספק קריאה כמותית עבור H2O2 מיטוכונדריאלי בתנאי גדילה רגילים ועם חשיפה לעקה חמצונית. פרוטוקול זה מסביר כיצד לייעל את תנאי ההדמיה באמצעות מערכת מיקרוסקופ קונפוקלי בדיסק מסתובב ולבצע ניתוח כמותי באמצעות תוכנה זמינה באופן חופשי. כלים אלה מאפשרים לאסוף מידע מרחבי-זמני עשיר על מיטוכונדריה הן בתוך תאים והן בקרב תאים באוכלוסייה. יתר על כן, זרימת העבודה המתוארת כאן יכולה לשמש לאימות ביו-חיישנים אחרים.
Introduction
מיטוכונדריה הם אברונים תאיים אאוקריוטים חיוניים הידועים בתפקידם בייצור ATP באמצעות זרחן חמצוני והובלת אלקטרונים1. בנוסף, מיטוכונדריה הם אתרים לאחסון סידן, סינתזה של שומנים, חומצות אמינו, חומצות שומן וצברי ברזל-גופרית, והעברת אותות 2,3. בתוך התאים, מיטוכונדריה יוצרים רשת דינמית עם מורפולוגיה ופיזור אופייניים, המשתנה בהתאם לסוג התא ולמצב המטבולי. יתר על כן, למרות שהמיטוכונדריה יכולים להתמזג ולהתחלק, לא כל המיטוכונדריה בתא שקולים. מחקרים רבים תיעדו את ההטרוגניות התפקודית של מיטוכונדריה בתוך תאים בודדים בתכונות כגון פוטנציאל ממברנה ומצב חמצוני 4,5,6. שינוי זה בתפקוד המיטוכונדריה נובע בחלקו מנזק לאברון כתוצאה ממוטציות mtDNA (המתרחשות בקצב גבוה יותר מאשר בדנ”א גרעיני) ומנזק חמצוני על ידי מיני חמצן תגובתי (ROS) הנוצרים בתוך אברוני 7,8,9 ומחוצה להם. הנזק לאברון מופחת על ידי מנגנוני בקרת איכות מיטוכונדריאליים שמתקנים את הנזק או מסלקים מיטוכונדריה שניזוקו מעבר לתיקון10.
מי חמצן (H 2 O2) הואמין חמצן תגובתי המהווה מקור לנזק חמצוני לחלבונים תאיים, חומצות גרעין ושומנים. עם זאת, H2 O2 משמש גם כמולקולת איתות המווסתת את הפעילות התאית באמצעות חמצון הפיך של תיולים בחלבוני מטרה11,12. H2O2 מופק מאלקטרונים הדולפים משרשרת הובלת האלקטרונים במיטוכונדריה ומאנזימים ספציפיים, כגון NADPH oxidase ומונואמין אוקסידאז 13,14,15,16,17,18,19,20. הוא גם מנוטרל על ידי מערכות נוגדות חמצון, כולל אלה המבוססות על תיורדוקסין וגלוטתיון21,22,23. לכן, ניתוח של רמות H 2 O2במיטוכונדריה הוא קריטי להבנת תפקידו של מטבוליט זה בתפקוד תקין של המיטוכונדריה והתאים ובתנאים של עקה חמצונית.
המטרה הכוללת של פרוטוקול זה היא לזהות H 2 O 2 מיטוכונדריאלי באמצעות ביו-חיישן מקודד גנטית H 2 O2, HyPer7, הממוקד לאברון (mtHyPer7). mtHyPer7 היא כימרה המורכבת מרצף האותות המיטוכונדריאלי מ-ATP9 (רצף Su9), צורה מעגלית של חלבון פלואורסצנטי ירוק (GFP), ותחום הקישור H 2 O2של חלבון OxyR מ-Neisseria meningitidis24 (איור 1). ב-GFP בעל תמורות מעגליות, ה-N וה-C-termini של GFP המקורי מתמזגים וטרמיני חדש נוצר ליד הכרומופור, מה שמקנה ניידות רבה יותר לחלבון ויכולת גדולה יותר של המאפיינים הספקטרליים שלו בהשוואה ל-GFP25 המקורי. האינטראקציה של תחום OxyR של mtHyPer7 עם H 2 O 2 היא בעלת זיקה גבוהה, H 2 O 2-סלקטיבית,ומובילה לחמצון הפיך של שאריות ציסטאין שמורות והיווצרות גשר דיסולפיד. שינויים קונפורמטיביים הקשורים לחמצון של OxyR מועברים ל-GFP בעל תמורות מעגליות ב-mtHyPer7, מה שגורם לשינוי ספקטרלי במקסימום העירור של כרומופור mtHyPer7 מ-405 ננומטר במצב מופחת ל-488 ננומטר במצב H 2 O2-oxidized26. לפיכך, היחס בין פלואורסצנטיות מ mtHyPer7 בתגובה לעירור ב 488 ננומטר לעומת 405 ננומטר משקף את חמצון הבדיקה על ידי H 2 O2.
באופן אידיאלי, ביו-חיישן אמור לספק קריאה כמותית מוחלטת בזמן אמת של מולקולת המטרה שלו. למרבה הצער, עם זאת, זה לא תמיד אפשרי במדידות בעולם האמיתי. במקרה של חיישני חמצון, כגון mtHyPer7, הקריאה בזמן אמת מושפעת מקצב ההפחתה של הגשר הדיסולפיד. מערכות החיזור המשמשות חיישנים ביולוגיים של ROS שונות, ואלה יכולות לשנות באופן דרמטי את דינמיקת תגובת הגשושית – כפי שניתן לראות בהשוואה בין HyPer7, מופחת על ידי מערכת thioredoxin, ו- roGFP2-Tsa2ΔCR, מופחת על ידי גלוטתיון בשמרים cytosol27. לפיכך, כדי להסיק מסקנה לגבי ריכוז H 2O2 יחסי mtHyPer7, יש להניח כי מערכת ההפחתה שומרת על קיבולת קבועה במהלך הניסוי. למרות שיקולים אלה, HyPer7 וגשושיות קשורות שימשו בהקשרים שונים להשגת מידע על H 2 O2 בתאים חיים25,28,29.
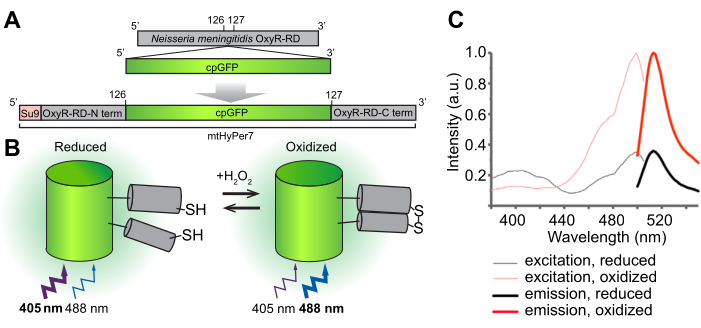
איור 1: תכנון, מנגנון מולקולרי וספקטרום עירור/פליטה של הביוסנסור H 2 O2 mtHyPer7. (A) הגשושית mtHyPer7 נגזרת על ידי החדרת GFP בעל תמורות מעגליות לתחום OxyR-RD מ-Neisseria meningitidis. הוא מכיל את רצף מיקוד המיטוכונדריה מתת-יחידה 9 של סינתאז ATP מ-Neurospora crassa (Su9). (B) איור של מנגנון החישה H 2 O2של mtHyPer7. חמצון של ציסטאין בתחום RD מגביר את הפליטה הפלואורסצנטית בעת עירור ב 488 ננומטר ומקטין את הפליטה הנוצרת על ידי עירור ב 405 ננומטר. (C) ספקטרום עירור ופליטה של HyPer7 בצורות מחומצנות ומופחתות. נתון זה הודפס מחדש באישור פאק ואחרים 24. קיצורים: GFP = חלבון פלואורסצנטי ירוק; cpGFP = GFP בעל תמורות מעגליות. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
הדמיה יחסימטרית זו של mtHyPer7 מציעה יתרונות חשובים לכימות H 2 O22 מיטוכונדריה 24,27; הוא מספק בקרה פנימית על ריכוז הבדיקה. בנוסף, השינוי בשיא העירור שנוצר על ידי חשיפה H 2 O 2אינו שלם, אפילו בריכוזים רוויים של H 2 O 2. לפיכך, הדמיית יחס יכולה להגביר את הרגישות על ידי שילוב שתי נקודות ספקטרליות בניתוח.
לגשושית mtHyPer7 המשמשת כאן יש זיקה גבוהה מאוד ל- H 2 O2ורגישות נמוכה יחסית ל- pH24, והיא ממוקדת בהצלחה ל- Caenorhabditis elegans mitochondria30. חלבון זה שימש גם בשמרים27,31. עם זאת, מחקרים קודמים הסתמכו על ביטוי פלסמיד של mtHyPer7, אשר גורם לשונות בין תא לתא בביטוי בדיקה27. בנוסף, המבנה המתואר בפרוטוקול זה שולב באזור שמור ונטול גנים על כרומוזום X32 באמצעות גישה מבוססת קריספר לאינטגרציה ללא סמנים. הביטוי של הגן הביוסנסורי המשולב נשלט גם על-ידי מקדם TEF1 החזק (איור 2). כתוצאה מכך, יש ביטוי יציב ועקבי יותר של הביו-סנסור באוכלוסיות תאי שמרים בהשוואה לזה שנצפה באמצעות ביטוי ביו-סנסור הנישא בפלסמיד, וניתן להפיץ תאים הנושאים את הביו-סנסור ללא צורך במדיה סלקטיבית.
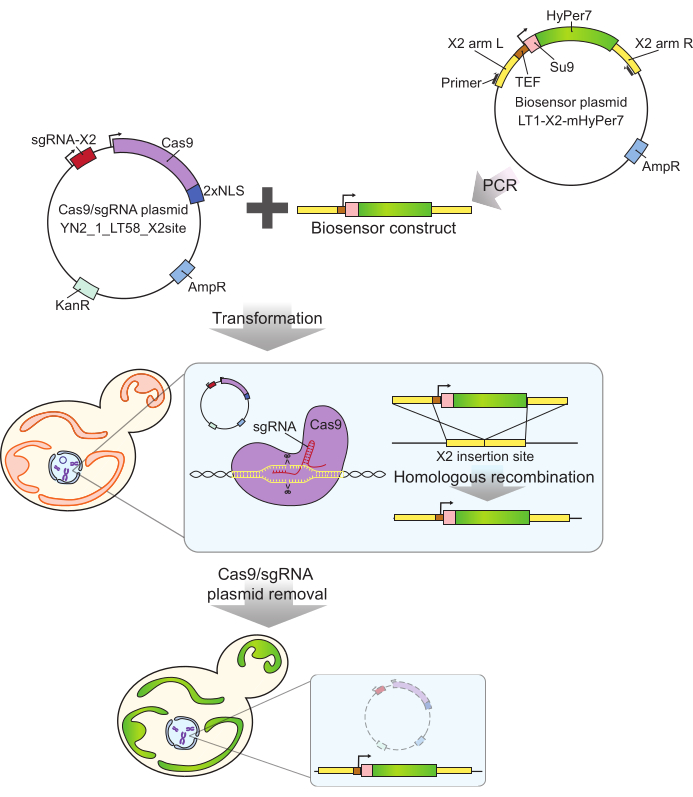
איור 2: יצירת תאים המבטאים mtHyPer7 על-ידי קריספר. הפלסמיד המכיל Cas9 ו- sgRNA (YN2_1_LT58_X2site) ומבנה הביוסנסור המכיל mtHyPer7 המוגבר ב- PCR מוכנסים לתאי שמרים ניצנים על ידי טרנספורמציית ליתיום אצטט. אתר ההחדרה נטול הגנים לכרומוזום X (X2) מזוהה ונחתך על ידי חלבון Cas9 עם sgRNA, והביוסנסור משולב בגנום על ידי רקומבינציה הומולוגית. לאחר זיהוי הטרנספורמטים המוצלחים על ידי סינון מיקרוסקופי, PCR מושבה וריצוף, פלסמיד Cas9 מוסר (נרפא) על ידי גידול במדיה לא סלקטיבית. קיצורים: sgRNA = RNA מדריך יחיד; TEF = גורם משפר תמלול. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
לבסוף, mtHyPer7 מציע יתרונות על פני biosensors אחרים ROS. לדוגמה, צבעים אורגניים המשמשים לזיהוי ROS (למשל, דיהידרואתידיום [DHE]2 ו- MitoSOX3) יכולים לייצר צביעה לא אחידה או לא ספציפית ולעתים קרובות מועברים בממסים כגון אתנול או דימתיל סולפוקסיד, הדורשים בקרות נוספות להשפעות ממס. סוג נוסף של ביו-חיישנים מסוג ROS הם ביו-חיישנים מבוססי העברת אנרגיה בתהודה פלואורסצנטית (FRET) (לדוגמה, Redoxfluor עבור מצב חמצון-חיזור תאי4, וחיישני מי חמצן HSP-FRET5, OxyFRET 6 ו-PerFRET6). בדיקות אלה מקודדות גנטית ורגישות מאוד באופן עקרוני וניתן לכוון אותן כמותית למיטוכונדריה באמצעות רצפי אותות מיטוכונדריאליים מאופיינים היטב. עם זאת, ישנם אתגרים בשימוש בבדיקות מבוססות FRET, כולל רקע עקב עירור צולב ודימום, ודרישות מחמירות לקרבה וכיוון של פלואורופורים עבור FRET להתרחש33,34. בנוסף, גשושיות FRET מורכבות משני חלבונים פלואורסצנטיים הדורשים מבנים גדולים יותר לביטוי בתאים בעלי עניין בהשוואה לגשושיות המזיזות ספקטרה. הפרוטוקול המתואר כאן פותח כדי לנצל את נקודות החוזק של הביו-חיישן מבוסס HyPer7, ולהשתמש בבדיקה קומפקטית, רציומטרית, בעלת זיקה גבוהה ומקודדת גנטית זו לצורך הדמיה כמותית של מי חמצן במיטוכונדריה בשמרים חיים.
Protocol
Representative Results
Discussion
בפרוטוקול זה, מתוארת שיטה לשימוש ב- mtHyPer7 כחיישן ביולוגי להערכת H 2 O2מיטוכונדריאלי בתאי שמרים ניצנים חיים. הביוסנסור נבנה בשיטה מבוססת קריספר ומוכנס לאזור שמור ללא גנים בגנום השמרים ללא שימוש בסמנים הניתנים לבחירה. בהשוואה לביו-חיישנים הנישאים בפלסמיד, אלה המשולבים באים לידי ביטוי בכל התאים וברמות עקביות, ומספקים תוצאות כימות אמינות יותר. לא נעשה שימוש בסמנים הניתנים לבחירה ליצירת תאים המבטאים mtHyPer7, מה שמאפשר שימוש במגוון רחב יותר של רקעי זן ומקל על השינוי הגנטי של תאים המבטאים ביוסנסורים. חלבון mtHyPer7 ממוקד נכון למיטוכונדריה ללא השפעות ניכרות על המורפולוגיה של המיטוכונדריה, תפקוד, פיזור או קצב גדילת תאים. mtHyPer7 מראה תגובה תלוית מינון לתוספת חיצונית H 2 O2. יתר על כן, mtHyPer7 מסוגל לדווח על ההטרוגניות של איכות המיטוכונדריה עם רזולוציה תת-תאית. לבסוף, שימוש במיקרוסקופ קונפוקלי של דיסק מסתובב, בניגוד למיקרוסקופ רחב שדה להדמיית חיישנים ביולוגיים ממוקדים במיטוכונדריה, גורם לפחות הלבנה של אור לפלואורופורים ומפיק תמונות ברזולוציה גבוהה לניתוח הבדלים תת-תאיים.
מגבלות וגישות חלופיות
שיטה זו אינה מתאימה לתאי הדמיה למשך יותר מ-10 דקות, מכיוון שהתאים יתייבשו מתחת לכיסוי. להדמיה לטווח ארוך יותר, עדיף להשתמש בשיטת אגר פד46 או לשתק תאים בצלחת תרבית תחתית זכוכית מלאה במדיום SC.
הבחירה של biosensor צריך להיות מונחה על ידי ריכוז המטרה בתנאי ניסוי. אם הרגישות של HyPer7 גבוהה מדי, מומלץ להשתמש בגרסת HyPer אחרת, כגון HyPer3 או HyPerRed47,48. עם זאת, יש לציין כי בדיקות HyPer אחרות רגישות יותר ל- pH. עבור רגישות גבוהה יותר, roGFP מבוסס peroxiredoxin, עשוי להיות מתאים יותר (roGFP2Tsa2ΔCR)27.
מצב החמצון היציב של חיישן H 2 O2קשור הן לקצב החמצון והן לקצב החיזור. קצב החמצון של חיישנים ביולוגיים נגרם בעיקר על ידי H 2 O2, אך קצב ההפחתה תלוי במערכות חיזור נוגדי חמצון הפעילות בתא ואברונים. הוכח כי HyPer7 מופחת בעיקר על ידי מערכת thioredoxin בציטוזול שמרים, וההפחתה שלו מהירה יותר מזו של roGFP2Tsa2ΔCR27. לכן, מנגנוני ההפחתה השונים ודינמיקת התגובה של הגשושית צריכים להילקח בחשבון בעת פירוש מדידות של חיישנים ביולוגייםH 2 O2. בפרט, כדי להסיק רמות H2 O2 מקריאת הביו-סנסור, יש להניח כי מערכת ההפחתה שומרת על קיבולת קבועה במהלך הניסוי. כחלופה לסקריפטים המתוארים כאן, מגוון תוכנות אחרות הועמדו לרשות הציבור באופן חופשי לניתוח חיישני חיזור49.
שלבים קריטיים
עם כל biosensor, זה קריטי כדי להוכיח כי biosensor עצמו אינו משפיע על התהליך הנמדד. לכן, השוואת הגידול והמורפולוגיה המיטוכונדריאלית של זנים תחת כל תנאי ניסוי חשובה. כאן, מורפולוגיה מיטוכונדריאלית מוערכת באמצעות MitoTracker Red, אשר מכתים מיטוכונדריה באופן תלוי פוטנציאל הממברנה. עם זאת, השוואה של המיטוכונדריה בתאים שלא עברו טרנספורמציה ועברו טרנספורמציה ביולוגית יכולה להתבצע על ידי צביעה עם טטרמתילרודאמין מתיל אסטר (TMRM), צבע חיוני מיטוכונדריאלי חלופי לחישת פוטנציאלים, או MitoTracker Green, אשר מכתים מיטוכונדריה ללא תלות בפוטנציאל הממברנה. אם יש חשד להשפעות מזיקות, הפחתת רמת הביטוי או שינוי אתר האינטגרציה עשויים לעזור.
אימות התנהגות המינון-תגובה של הגשושית ויחס האות לרעש של טכניקת ההדמיה חיוניים גם הם לאיסוף תוצאות חזקות. אם השונות בתוך קבוצה עולה על השונות בין קבוצות, קשה לזהות את ההבדלים. שונות תוך-קבוצתית עשויה לנבוע משונות אמיתית באוכלוסייה, או מרעש בתהליך האיתור. השלבים העיקריים להגדלת יחס האות:רעש הם רכישת תמונה (טווח ערכי פיקסלים ורעש), חיסור רקע וסף.
ניתן גם להפחית את השפעות הרעש במהלך שלבי החישוב. הגישה הפשוטה ביותר היא לחשב את העוצמה הממוצעת המשוקללת ממדידות תמונת היחס (תוצאות.csv), כאשר כל פיקסל מייצג את היחס המקומי בין יעילות העירור. זה מייצר יחס “פיקסלים”. עם זאת, אם יחס אות התמונה:רעש נמוך, ניתן לקבל תוצאות חזקות יותר על ידי חישוב העוצמה הממוצעת המשוקללת עבור החזר השקעה הן בערוצי המונה והן בערוץ המכנה, ולאחר מכן חישוב היחס בין שני אמצעים משוקללים אלה (יחס “אזורי”).
לבחירת שיטת סף, הפקודה פיג’י תמונה | התאמה | ניתן להשתמש בסף אוטומטי כדי לנסות באופן אוטומטי את כל שיטות פיג’י המובנות. כדי להעריך סגמנטציה (סף), מסיכה שנשמרה מומרת לבחירה על ידי לחיצה על עריכה | בחירה | צור בחירה, נוסף למנהל החזר ההשקעה (על-ידי הקשה על T) ולאחר מכן הופעל בקובץ התמונה הגולמית. אם המיטוכונדריה אינם מזוהים כראוי, יש לנסות שיטת פילוח אחרת.
בעת השוואת תמונות, חיוני לרכוש את כל התמונות עם תנאי הדמיה זהים, כמו גם להציג את כל התמונות עם שיפור ניגודיות זהה.
יש לקחת בחשבון את התנועה המיטוכונדריאלית בעת מיטוב תנאי ההדמיה. אם המיטוכונדריה נעים באופן משמעותי בין עירור של 405 ל-488 ננומטר, תמונת היחס לא תהיה מדויקת. מומלץ לשמור על זמן חשיפה <500 אלפיות השנייה ולשנות עירור בשיטה המהירה ביותר הקיימת (למשל דופק טריגר או מסנן כוונון אקוסטי-אופטי). בעת לכידת מחסנית Z, יש לבצע את שני העירורים עבור כל צעד Z לפני המעבר לשלב Z הבא.
לצורך הצגת התוצאות, שינויים בגוון (צבע) ברורים יותר לעין האנושית מאשר שינויים בעוצמה. לכן, ערך היחס מומר לסרגל צבעים לפרשנות חזותית קלה יותר. ניתן לבטל אפנון של תמונות צבעוניות, שבהן כל הפיקסלים במיטוכונדריה מופיעים באותה בהירות, או עם אפנון עוצמה, שבו עוצמת הפיקסלים בתמונה המקורית משמשת לקביעת עוצמות בתמונה הצבעונית.
שינוי ופתרון בעיות
כחלופה לאישור תפקוד המיטוכונדריה על ידי אתגר עם פרקוואט, תאים עשויים להיות מצופים העתק או מחוסנים למקורות פחמן מותססים ולא מותססים.
לחיסור רקע, חיסור כדור מתגלגל (על-ידי ניווט אל תהליך | חיסור רקע…) ניתן להשתמש גם כדי להסיר אי-אחידות תאורה. יש לוודא כי נוכחות התאים אינה משנה את הרקע המופחת (על ידי סימון האפשרות ליצור רקע ובחינת התוצאה).
לסיכום, הגשושית mtHyPer7 מספקת שיטה עקבית וזעיר פולשנית לקשר את המצב המורפולוגי והתפקודי של מיטוכונדריית שמרים בתאים חיים, ומאפשרת לחקור גורם עקה תאי חשוב ומולקולת איתות במערכת מודל גנטית נגישה ונגישה.
Disclosures
The authors have nothing to disclose.
Acknowledgements
המחברים מודים לקתרין פילפו לופז על סיוע טכני מומחה. עבודה זו נתמכה על ידי מענקים מהמכונים הלאומיים לבריאות (NIH) (GM122589 ו-AG051047) ל-LP.
מחקרים אלה השתמשו במשאב המשותף Confocal and Specialized Microscopy של מרכז הסרטן המקיף ע”ש הרברט אירווינג באוניברסיטת קולומביה, שמומן בחלקו באמצעות מענק התמיכה של מרכז הסרטן NIH/NCI P30CA013696.
Materials
| 100x/1.45 Plan Apo Lambda objective lens | Nikon | MRD01905 | |
| Adenine sulfate | Sigma-Aldrich | #A9126 | |
| Bacto Agar | BD Difco | #DF0145170 | |
| Bacto Peptone | BD Difco | #DF0118170 | |
| Bacto Tryptone | BD Difco | #DF211705 | |
| Bacto Yeast Extract | BD Difco | #DF0127179 | |
| BamHI | New England Biolabs | R0136S | |
| BglII | New England Biolabs | R0144S | |
| Carbenicilin | Sigma-Aldrich | C1389 | |
| Carl Zeiss Immersol Immersion Oil | Carl Zeiss | 444960 | |
| Dextrose (D-(+)-Glucose) | Sigma-Aldrich | #G8270 | |
| E. cloni 10G chemical competent cell | Bioserch Technologies | 60108 | |
| FIJI | NIH | Schindelin et al 2012 | |
| G418 (Geneticin) | Sigma-Aldrich | A1720 | |
| GFP emission filter | Chroma | 525/50 | |
| Gibson assembly | New England Biolabs | E2611 | |
| Graphpad Prism 7 | GraphPad | https://www.graphpad.com/scientific-software/prism/ | |
| H2O2 (stable) | Sigma-Aldrich | H1009 | |
| HO-pGPD-mito-roGFP-KanMX6-HO | Pon Lab | JYE057/EP41 | Liao et al 20201 |
| Incubator Shaker | New Brunswick Scientific | E24 | |
| KAPA HiFi PCR kit | Roche Sequencing and Life Science, Kapa Biosystems, Wilmington, MA | KK1006 | |
| L-arginine hydrochloride | Sigma-Aldrich | #A8094 | |
| laser | Agilent | 405 and 488 nm | |
| L-histidine hydrochloride | Sigma-Aldrich | #H5659 | |
| L-leucine | Sigma-Aldrich | #L8912 | |
| L-lysine hydrochloride | Sigma-Aldrich | #L8662 | |
| L-methionine | Sigma-Aldrich | #M9625 | |
| L-phenylalanine | Sigma-Aldrich | #P5482 | |
| L-tryptophan | Sigma-Aldrich | #T8941 | |
| L-tyrosine | Sigma-Aldrich | #T8566 | |
| mHyPer7 plasmid | This study | JYE116 | |
| Microscope coverslips | ThermoScientific | 3406 | #1.5 (170 µm thickness) |
| Microscope slides | ThermoScientific | 3050 | |
| MitoTracker Red CM-H2Xros | ThermoFisherScientific | M7513 | |
| NaCl | Sigma-Aldrich | S9888 | |
| NEBuilder HiFi Assembly Master Mix | New England Biolabs | E2621 | |
| Nikon Elements | Nikon | Microscope acquisition software | |
| Nikon Ti Eclipse inverted microscope | Nikon | ||
| Paraquat (Methyl viologen dichloride hydrate) | Sigma-Aldrich | Cat. #856177 | |
| RStudio | Posit.co | Free desktop version | |
| Spectrophotometer | Beckman | BU530 | |
| Stagetop incubator | Tokai Hit | INU | |
| Uracil | Sigma-Aldrich | #U1128 | |
| Yeast nitrogen base (YNB) containing ammonium sulfate without amino acids | BD Difco | #DF0919073 | |
| YN2_1_LT58_X2site | Addgene | 177705 | Pianale et al 2021 |
| Zyla 4.2 sCMOS camera | Andor |
References
- vander Bliek, A. M., Sedensky, M. M., Morgan, P. G. Cell biology of the mitochondrion. Genetics. 207 (3), 843-871 (2017).
- McBride, H. M., Neuspiel, M., Wasiak, S. Mitochondria: more than just a powerhouse. Current Biology. 16 (14), 551-560 (2006).
- Shi, R., Hou, W., Wang, Z. -. Q., Xu, X. Biogenesis of iron-sulfur clusters and their role in DNA metabolism. Frontiers in Cell and Developmental Biology. 9, 735678 (2021).
- McFaline-Figueroa, J. R., et al. Mitochondrial quality control during inheritance is associated with lifespan and mother-daughter age asymmetry in budding yeast. Aging Cell. 10 (5), 885-895 (2011).
- Higuchi-Sanabria, R., et al. Mitochondrial anchorage and fusion contribute to mitochondrial inheritance and quality control in the budding yeast Saccharomyces cerevisiae. Molecular Biology of the Cell. 27 (5), 776-787 (2016).
- Higuchi-Sanabria, R., et al. Role of asymmetric cell division in lifespan control in Saccharomyces cerevisiae. FEMS Yeast Research. 14 (8), 1133-1146 (2014).
- Lam, Y. T., Aung-Htut, M. T., Lim, Y. L., Yang, H., Dawes, I. W. Changes in reactive oxygen species begin early during replicative aging of Saccharomyces cerevisiae cells. Free Radical Biology & Medicine. 50 (8), 963-970 (2011).
- Laun, P., et al. Aged mother cells of Saccharomyces cerevisiae show markers of oxidative stress and apoptosis. Molecular Microbiology. 39 (5), 1166-1173 (2001).
- Doudican, N. A., Song, B., Shadel, G. S., Doetsch, P. W. Oxidative DNA damage causes mitochondrial genomic instability in Saccharomyces cerevisiae. Molecular and Cellular Biology. 25 (12), 5196-5204 (2005).
- Roca-Portoles, A., Tait, S. W. G. Mitochondrial quality control: from molecule to organelle. Cellular and Molecular Life Sciences. 78 (8), 3853-3866 (2021).
- Sies, H., Berndt, C., Jones, D. P. Oxidative stress. Annual Review of Biochemistry. 86, 715-748 (2017).
- Sies, H., Jones, D. P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nature Reviews Molecular Cell Biology. 21 (7), 363-383 (2020).
- Imlay, J. A., Fridovich, I. Assay of metabolic superoxide production in Escherichia coli. The Journal of Biological Chemistry. 266 (11), 6957-6965 (1991).
- Fridovich, I. Mitochondria: are they the seat of senescence. Aging Cell. 3 (1), 13-16 (2004).
- Quinlan, C. L., Perevoshchikova, I. V., Hey-Mogensen, M., Orr, A. L., Brand, M. D. Sites of reactive oxygen species generation by mitochondria oxidizing different substrates. Redox Biology. 1 (1), 304-312 (2013).
- Griendling, K. K., Minieri, C. A., Ollerenshaw, J. D., Alexander, R. W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circulation Research. 74 (6), 1141-1148 (1994).
- Griendling, K. K., Sorescu, D., Ushio-Fukai, M. NAD(P)H oxidase: role in cardiovascular biology and disease. Circulation Research. 86 (5), 494-501 (2000).
- Edmondson, D. E., Binda, C., Wang, J., Upadhyay, A. K., Mattevi, A. Molecular and mechanistic properties of the membrane-bound mitochondrial monoamine oxidases. Biochemistry. 48 (20), 4220-4230 (2009).
- Ramsay, R. R., Singer, T. P. The kinetic mechanisms of monoamine oxidases A and B. Biochemical Society Transactions. 19 (1), 219-223 (1991).
- Ramsay, R. R. Kinetic mechanism of monoamine oxidase A. Biochemistry. 30 (18), 4624-4629 (1991).
- Handy, D. E., Loscalzo, J. Redox regulation of mitochondrial function. Antioxidants & Redox Signaling. 16 (11), 1323-1367 (2012).
- Wood, Z. A., Schröder, E., Robin Harris, J., Poole, L. B. Structure, mechanism and regulation of peroxiredoxins. Trends in Biochemical Sciences. 28 (1), 32-40 (2003).
- Slade, L., et al. Examination of the superoxide/hydrogen peroxide forming and quenching potential of mouse liver mitochondria. Biochimica et Biophysica Acta. General Subjects. 1861 (8), 1960-1969 (2017).
- Pak, V. V., et al. Ultrasensitive genetically encoded indicator for hydrogen peroxide identifies roles for the oxidant in cell migration and mitochondrial function. Cell Metabolism. 31 (3), 642-653 (2020).
- Topell, S., Hennecke, J., Glockshuber, R. Circularly permuted variants of the green fluorescent protein. FEBS Letters. 457 (2), 283-289 (1999).
- Belousov, V. V., et al. Genetically encoded fluorescent indicator for intracellular hydrogen peroxide. Nature Methods. 3 (4), 281-286 (2006).
- Kritsiligkou, P., Shen, T. K., Dick, T. P. A comparison of Prx- and OxyR-based H2O2 probes expressed in S. cerevisiae. The Journal of Biological Chemistry. 297 (1), 100866 (2021).
- Baird, G. S., Zacharias, D. A., Tsien, R. Y. Circular permutation and receptor insertion within green fluorescent proteins. Proceedings of the National Academy of Sciences. 96 (20), 11241-11246 (1999).
- Abedi, M. R., Caponigro, G., Kamb, A. Green fluorescent protein as a scaffold for intracellular presentation of peptides. Nucleic Acids Research. 26 (2), 623-630 (1998).
- Onukwufor, J. O., et al. A reversible mitochondrial complex I thiol switch mediates hypoxic avoidance behavior in C. elegans. Nature Communications. 13 (1), 2403 (2022).
- Vega, M., et al. Antagonistic effects of mitochondrial matrix and intermembrane space proteases on yeast aging. BMC Biology. 20 (1), 160 (2022).
- Torello Pianale, L., Rugbjerg, P., Olsson, L. Real-time monitoring of the yeast intracellular state during bioprocesses with a toolbox of biosensors. Frontiers in Microbiology. 12, 802169 (2022).
- Imani, M., Mohajeri, N., Rastegar, M., Zarghami, N. Recent advances in FRET-based biosensors for biomedical applications. Analytical Biochemistry. 630, 114323 (2021).
- Zadran, S., et al. Fluorescence resonance energy transfer (FRET)-based biosensors: visualizing cellular dynamics and bioenergetics. Applied Microbiology and Biotechnology. 96 (4), 895-902 (2012).
- Gietz, R. D., Woods, R. A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods in Enzymology. 350, 87-96 (2002).
- Liao, P. -. C., Wolken, D. M. A., Serrano, E., Srivastava, P., Pon, L. A. Mitochondria-associated degradation pathway (MAD) function beyond the outer membrane. Cell Reports. 32 (2), 107902 (2020).
- Higuchi-Sanabria, R., Swayne, T. C., Boldogh, I. R., Pon, L. A. Live-cell imaging of mitochondria and the actin cytoskeleton in budding yeast. Methods in Molecular Biology. 1365, 25-62 (2016).
- Liao, P. -. C., Yang, E. J., Pon, L. A. Live-cell imaging of mitochondrial redox state in yeast cells. STAR Protocols. 1 (3), 100160 (2020).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Collins, T. J. ImageJ for microscopy. BioTechniques. 43, 25-30 (2007).
- Chazotte, B. Labeling mitochondria with MitoTracker dyes. Cold Spring Harbor Protocols. 2011 (8), 990-992 (2011).
- Aguilaniu, H., Gustafsson, L., Rigoulet, M., Nyström, T. Asymmetric inheritance of oxidatively damaged proteins during cytokinesis. Science. 299 (5613), 1751-1753 (2003).
- Erjavec, N., Larsson, L., Grantham, J., Nyström, T. Accelerated aging and failure to segregate damaged proteins in Sir2 mutants can be suppressed by overproducing the protein aggregation-remodeling factor Hsp104p. Genes & Development. 21 (19), 2410-2421 (2007).
- Erjavec, N., Cvijovic, M., Klipp, E., Nyström, T. Selective benefits of damage partitioning in unicellular systems and its effects on aging. Proceedings of the National Academy of Sciences. 105 (48), 18764-18769 (2008).
- Erjavec, N., Nyström, T. Sir2p-dependent protein segregation gives rise to a superior reactive oxygen species management in the progeny of Saccharomyces cerevisiae. Proceedings of the National Academy of Sciences. 104 (26), 10877-10881 (2007).
- Davidson, R., Liu, Y., Gerien, K. S., Wu, J. Q. Real-time visualization and quantification of contractile ring proteins in single living cells. Methods in Molecular Biology. 1369, 9-23 (2016).
- Bilan, D. S., et al. HyPer-3: a genetically encoded H2O2 probe with improved performance for ratiometric and fluorescence lifetime imaging. ACS Chemical Biology. 8 (3), 535-542 (2013).
- Ermakova, Y. G., et al. Red fluorescent genetically encoded indicator for intracellular hydrogen peroxide. Nature Communications. 5 (1), 5222 (2014).
- Fricker, M. D. Quantitative redox imaging software. Antioxidants & Redox Signaling. 24 (13), 752-762 (2016).
- Yang, E. J., Pernice, W. M., Pon, L. A. A role for cell polarity in lifespan and mitochondrial quality control in the budding yeast Saccharomyces cerevisiae. iSCIENCE. 25 (3), 103957 (2022).
