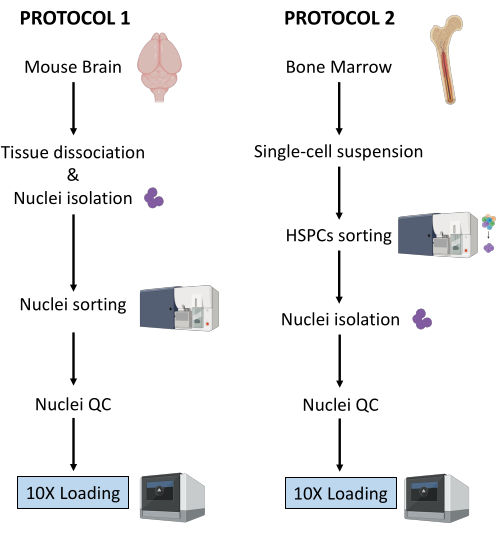
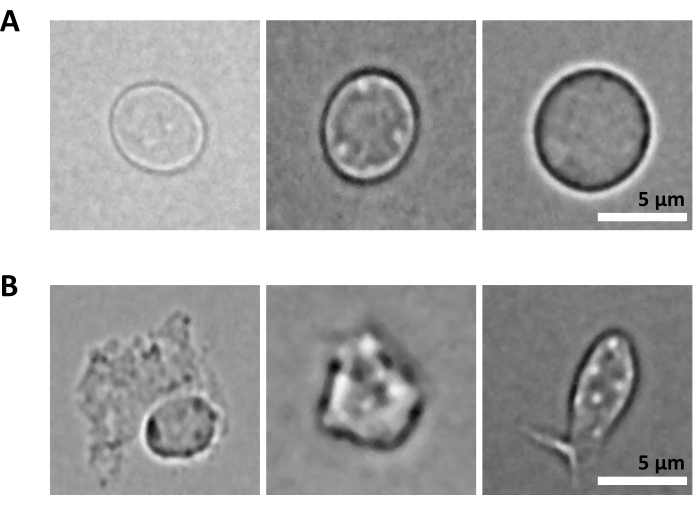
Eksperimentelle prosedyrer ble utført under streng regulatorisk overholdelse av protokoller godkjent av Komiteen for etikk i dyreforsøk (CETEA). For hjernekjerneisolasjonen ble 3 måneder gamle C57BL/6-mus brukt. Til benmargsisoleringen ble det brukt 8 uker gamle kvinnelige C57BL/6J-mus som veide 18 g. 1. Rensing av kjerner fra musehjernen MERK: Bruk lateks- eller nitrilhansker til enhver tid under prosedyren. Det anbefales på det sterkeste å ha to personer som utfører eksperimentet, å ha trinn 1 til 3 (dvs. fremstilling av enkeltkjernesuspensjonen) utført av en person, og trinn 4 (dvs. forberedelse av sortereren) utført parallelt av en annen person. Siden protokollen er svært tidssensitiv, er det avgjørende å minimere prøvebehandlingstiden ved å ha sortereren klar så snart enkeltkjernesuspensjonen er forberedt. Fremstilling av reagenser og materialerSteriliser disseksjonsverktøyene forsiktig med autoklav (ved 121 °C i 20 minutter) og vask dem med etanol 70 % rett før bruk. Forbered en petriskål per prøve, fylt med 2-3 ml iskald 1x Dulbeccos fosfatbufret saltvann (DPBS). Avkjøl mikrosentrifugen til 4 °C, fyll en bøtte med is og legg glassdounce-homogenisatoren på is. Forbered kjernelysisbuffer ved å tilsette digitonin for en endelig konsentrasjon på 0,01%, 10 ml per prøve. Forbered fargebufferen ved å tilsette RNase-hemmere til cellefargebufferen for en endelig konsentrasjon på 0,2 U / μL, 20 ml per prøve. Forbered DPBS 0,04% BSA ved å tilsette RNase-hemmere for en endelig konsentrasjon på 0,2 U / μL, 2 ml per prøve. Forbered 1 ml fortynnet kjernebuffer i henhold til Multiome-protokollen25. Oppbevar alle reagenser og prøver på is. Vev disseksjonOfre mus ved hjelp av protokoller godkjent av institusjonen. I denne protokollen ble musene halshugget etter en ketamin / xylazin overdose. Klipp musehodet med saks og fjern hjernen fra skallen som beskrevet i Meyerhoff et al.26. Overfør umiddelbart hjernen til en petriskål tilberedt med den iskalde 1x DPBS under et lysemitterende diode (LED)-opplyst stereomikroskop. Klipp hjernevevet med en skalpell for å skille hjerneområder av interesse (f.eks. Entorhinal cortex, hippocampus, prefrontal cortex) og overfør hver region til en egen petriskål som inneholder iskald 1x DPBS. Hold på is. Hakk vevet i <0,5 cm stykker med en skalpell for å lette homogeniseringen i det følgende trinnet. Med en P1000-mikropipette overfører du hakket vev og 1x DPBS fra petriskålen til et 1,5 ml rør. Sørg for å bruke rør laget av protein-lavbindende plast. La vevstykkene skille seg ved tyngdekraften. Fjern forsiktig overskuddet av 1x DPBS ved hjelp av en P1000 mikropipette.MERK: Etter dette trinnet er det mulig å knipse og fryse det hakkede vevet ved å overføre proteinets lavbindende rør til tørris og deretter lagre ved -80 °C til du fortsetter med kjerneisolering. Kjerner isolasjonFyll glassdunken med 2 ml iskald kjernelysbuffer med 0,01 % digitonin. Tilsett vevsbitene i spretten.MERK: Hvis du arbeider med ferskfrosset vev, tilsett det hakkede frosne vevet direkte til kjernelysebufferen 0,01% digitonin; Ikke la vevet tine før. Homogeniser ved hjelp av et glass dounce vev homogenisator 25 ganger med pestle A og deretter 25 ganger med pestle B. Overfør homogenatet til et 15 ml rør. Legg til ytterligere 2 ml iskald kjernelysbuffer med 0,01% digitonin og inkuber på is i 5 minutter. Sentrifugekjerner ved 500 x g i 5 minutter ved 4 °C. Fjern supernatanten med en mikropipette og tilsett 4 ml iskald kjernelysbuffer med 0,01 % digitonin. Inkuber på is i 5 min og filtrer gjennom en 40 μm cellesil. Sentrifugekjerner ved 500 x g i 5 minutter ved 4 °C og fjern supernatanten med en mikropipette. Tilsett 4 ml fargebuffer for å vaske kjernene og sentrifuger ved 500 x g i 5 minutter ved 4 °C. Fjern supernatanten med en mikropipette og resuspender pelleten i 4 ml fargebuffer. Filtrer gjennom en 40 μm cellesil og sentrifuge ved 500 x g i 5 minutter ved 4 °C. Resuspender i 1 ml PBS med 0,04 % BSA. Telle kjerner for å sikre konsistens av vev / kjerner forberedelse på tvers av ulike prøver. Det forventes å oppnå lignende kjernetall fra de samme hjernegruppene:Tilsett 10 μL 0,4% trypanblå til et tomt 0,5 ml rør. Tilsett 10 μL av kjernene og bland 5x ved pipettering. Tell kjerner ved hjelp av en automatisert celleteller etter leverandørens anbefalinger. Hold kjernene på is. Forbered kjerner for sortering.MERK: De ekstraherte kjernene inneholder 7-AAD, og denne fargingen brukes til rensing av fluorescensaktivert cellesortering (FACS).Overfør 100 μL kjerner til et FACS-rør for ufarget kontroll. Tilsett 10 μL 7-AAD til de gjenværende kjernene, og hold 5 minutter ved 4 °C. Sorter minimum 0,5 x 106 kjerner etter FACS for å eliminere dubletter og rusk. Kjernesortering ved hjelp av en FACSMERK: Mens kjernesortering kan utføres på et bredt utvalg av cellesorterere, er prosedyren for bruk av instrumentene BD FACSAria Fusion eller BD FACSAria III beskrevet her. Det anbefales sterkt at kalibreringen og oppsettet av cellesortereren utføres under tilsyn, eller av en erfaren bruker av instrumentet. For å redusere prøvebehandlingstiden er det avgjørende å ha sortereren klar så snart den enkle kjernesuspensjonen er klargjort.Kalibrering av FACS-instrumentSlå på cellesorteringen og datamaskinen. Når programvaren er koblet til instrumentet, starter du den fluidiske oppstartsprosedyren. Velg Cytometer > Fluidic oppstart i hovedmenyen og følg de fire trinnene. Klikk på Ferdig etter å ha fullført hver. Sett inn 70 μm dysen, slå på strømmen og la strømmen stabilisere seg i 15 minutter. Juster amplitude for å få dråpeformasjon og klikk på Sweet Spot. Sett filteret med nøytral tetthet (N.D) 1.0 og åpne grensesnittet for cytometeroppsett og sporing (CST). Daglig kvalitetskontroll: Fortynn CST-perler i FACS-medium (se leverandørens anbefalinger) og utfør CST-kontroll. Når du er ferdig, erstatter du N.D 1.0 med N.D 2.0. Fortynn Accudrops i FACS-medium (se leverandørens anbefalinger) og utfør dråpeforsinkelse som beskrevet i trinn 6 til 10. Velg Accudrop Drop Delay-eksperimentet i eksperimentmalen og åpne sorteringsoppsettet for røret. Inne i det nedre kameravinduet klikker du på Spenning og deretter på Optisk filter for å aktivere ladning på avbøyningsplatene og bruke et spesifikt optisk filter foran kameraet. Sørg for at kvadranten på høyre side indikerer 100. Juster om nødvendig den røde laserskruen for å optimalisere laserpåvirkningen. Juster strømningshastigheten for å nå hastigheten på 1 000 til 3 000 hendelser per sekund. Klikk på Sorter og avbryt. Sørg for at venstre kvadrant er lik 100, og høyre kvadrant er 0. Hvis venstre kvadrant er under 95, utfør automatisk forsinkelse. Klikk på Spenning, deretter Test Sorter. Kontroller kvaliteten på sidestrømmene som legges i oppsamlingsrørene. Juster om nødvendig posisjonen til sidestrømmene ved å flytte glidebryterne. Oppsett av FACS-instrument for kjernesortering.Start oppkjøpet av de ufargede kjernene. Disse brukes til å definere forover- og sidespredningene, og detektorspenningen for 7-ADD-parameteren. Still inn parametrene slik at 7-AAD-signalet til den ufargede prøven faller innenfor det første tiåret av loggskalaen på punktplottet. Begynn å anskaffe røret av 7-AAD-fargede kjerner og definer kjernepopulasjonene ved å bruke en gating-strategi basert på (1) FSC-A / SCC-A og deretter FSC-H / SSC-H for størrelse og granularitet, (2) FSC-H / FSC-A for dublettdiskriminering, og (3) SSC-A / 7-AAD for 7-AAD positive kjerner (se figur 2A). Forsikre deg om at strømmen og nedbøyningen er stabile. I sidestrømkameraet slår du på testsorteringen, spenning PÅ, og bekrefter nøyaktig fallsortering i et 1,5 ml rør montert på venstre side. I vinduet Sorteringsoppsett velger du interessegruppen, som definert i trinn 2 (ovenfor). I Målhendelser velger du terskelen i Kontinuerlig for å få minst 0,5 x 106 kjerner per prøve. Under Presisjon velger du 4-veis renhet. Når du er klar, klikker du på Sorter og OK for å starte med kjernesortering. Kvalitetskontroll og telling av rensede kjernerMERK: Dette trinnet skal bare gjøres under piloteksperimentet for optimalisering av prøveprepareringstrinn, med mål om å teste renheten til de sorterte kjernene som skal lastes på 10X krombrikken. Når protokollen er fullstendig optimalisert, anbefales det ikke å utføre dette kvalitetskontrolltrinnet i oppfølgingsforsøkene for å unngå unødvendig sløsing med innsamlede kjerner som kan være tilgjengelige i lave antall.Renhetskontroll ved flowcytometriOverfør 10 μL av de sorterte kjernene til et nytt FACS-rør som inneholder 90 μL DPBS med 2% varmeinaktivert Fetal Bovine Serum (HI-FBS). Innhent og registrer data etter sortering for å bekrefte sorteringsrenhet og levedyktighet. Sørg for at minst 98% av kjernene vises i porten av interesse, som definert i 4.2 (se figur 2B). Teller de rensede kjerneneSentrifugesorterte kjerner i 5 minutter ved 500 x g og ved 4 °C og fjern supernatanten forsiktig helt ved hjelp av en mikropipette. Resuspender i 100 μL fortynnet kjernebuffer. Tilsett 10 μL 0,4% trypanblå til et tomt 0,5 ml rør. Tilsett 10 μL av de sorterte kjernene og bland 5x ved pipettering. Tell kjerner ved hjelp av en automatisert celleteller etter leverandørens anbefalinger. Juster kjernekonsentrasjonen til 3,5 x 106/ml, dvs. 16 000 kjerner per 5 μL. Kvalitetskontroll av rensede kjerner ved mikroskopiMERK: Dette trinnet skal bare gjøres under piloteksperimentet for optimalisering av prøveprepareringstrinn for å teste kvaliteten på kjernene som skal lastes på 10X krombrikken. Når protokollen er fullstendig optimalisert, anbefales det ikke å utføre dette kvalitetskontrolltrinnet i oppfølgingsforsøkene for å unngå unødvendig sløsing med innsamlede kjerner som kan være tilgjengelige i lave antall.Sørg for at lysbilder med mikroskop og dekselslipp er rene og støvfrie. Hvis nødvendig, vask og skyll dekslene med absolutt etanol og tørk dem med lofrie kluter. Fordel 25 μL poly-l-lysin i glidebrønnene som skal brukes og inkuber i 10 minutter ved romtemperatur (RT), beskyttet mot støv. Fjern overflødig poly-l-lysin og tilsett 10 μL av den rensede kjernesuspensjonen. Inkubere i 5 min ved RT, beskyttet mot støv. Tilsett en dråpe monteringsmedium til hver brønn, unngå bobler. Legg en dekkslipp på toppen av de frøede brønnene. Dekk til med papirservietter og trykk dekselet hardt for å fjerne overflødig monteringsmedium. Vær forsiktig så du ikke flytter dekselet, og ikke rengjør overflødig monteringsmedium. Ta flere bilder med et invertert mikroskop med lysfelt og en minimumsforstørrelse på 40x. Utfør multiomanalyse.Fortsett umiddelbart til Chromium Next GEM Single Cell Multiome ATAC + Gene Expression User Guide (CG000338 – Rev F)25. 2. Rensing av kjerner fra mus, benmarg, hematopoietisk stamme og stamceller (HSPCs) MERK Denne protokollen beskriver rensing av kjerner fra tre undergrupper av benmargen HSPCs: lineage-c-Kit + Sca-1 + hematopoietiske stamceller (HSC), avstamning-c-Kit + Sca-1-CD34 + FcγR- vanlige myeloide forfedre (CMP), og avstamning-c-Kit + Sca-1-CD34 + FcγR + granulocyt-monocytprogenitorer (GMP). Bruk latex eller nitrilhansker til enhver tid under prosedyren. Denne protokollen er en tilpasning av 10X Genomics Demonstrated Protocol – Nuclei Isolation for Single Cell Multiome ATAC + GEX sekvensering (CG000365 – Rev C) 27. Modifikasjoner er introdusert i den opprinnelige protokollen for å maksimere kjerneutvinning. Det anbefales på det sterkeste å ha to personer som utfører eksperimentet, for å ha trinn 1. til 3. (dvs. fremstilling av encelleoppløsningen) utført av en person, og trinn 4 (dvs. fremstilling av sorteringen) utført parallelt av en annen person. Siden protokollen er svært tidssensitiv, er det avgjørende å minimere prøvebehandlingstiden ved å ha sortereren klar så snart encellesuspensjonen er klargjort. Fremstilling av reagenser og materialerFyll en bøtte med is. Forbered FACS-bufferen: DPBS med 2% HI-FBS-løsning (ca. 500 ml for 6 prøver) og filtrer gjennom et 0,2 μm filter. Forbered oppsamlingsmediet: DPBS med 10% HI-FBS-løsning (500 μL per prøve), og filtrer gjennom et 0,2 μm filter. Isolering av benmargcellerOfre mus ved hjelp av protokoller godkjent av institusjonen. I dette forsøket ble musene ofret av cervikal dislokasjon etter en ketamin / xylazin overdose. Spray musens mage og bakben med 70% etanol. Bruk steril tang og saks til å lage et lite snitt midt i underlivet og åpne bukhinnen fra bakbeina til mellomgulvet (tilleggsfigur 1). Lag et ekstra kutt for hvert bakben vinkelrett på den åpnede bukhinnen, ta deretter tak i hver side av et av disse ekstra kuttene og trekk det fra hverandre for å skrelle av huden fra begge bakbenene forbi ankelleddet for å eksponere musklene i begge bakbenene (tilleggsfigur 1A). Linje saksen langs ryggraden ved hofteleddet på det ene bakbenet for å skjære ut benet uten å skjære gjennom lårbenet (tilleggsfigur 1B, C). Gjenta det samme for det andre benet. For å isolere lårbenet, skjær det meste av muskelvevet ut, og hold deretter lårbenet og tibia i hver hånd med fingertuppene i leddet (tilleggsfigur 1D, E). Brett benet forsiktig mot den naturlige bøyningen for å forskyve tibia fra lårbenet (tilleggsfigur 1E) og klipp deretter bindevevet forsiktig med saks for å skille lårben og tibia. Bruk saksen med lette vridningsbevegelser for å forskyve biten av ryggraden fra den øverste enden av lårbenet (tilleggsfigur 1E). Rengjør det isolerte lårbenet med silkepapir for å fjerne gjenværende muskel og bindevev. Hold kaldt på is i en 12-brønns platebrønn fylt med 2 ml DMEM (1x) + GlutaMAX-I. Når alle lårbenene er samlet, må du sørge for at muskler og fibrøst vev er helt fjernet fra beinet. Ikke kutt benet åpent for å (a) holde margen inne steril og (b) unngå å miste celler i brønnen. Bruk følgende trinn for å skylle cellene fra to lårben på en mus, tilpasset fra Haag og Murthy28. Forbered ett 1,5 ml og ett 0,5 ml rør. Tilsett 150 μL av FACS-bufferen til 1,5 ml røret, stikk deretter et hull i bunnen av 0,5 ml røret ved hjelp av en 18 G nål og sett 0,5 ml røret inn i 1,5 ml røret. Åpne den distale delen av hvert lårben med kirurgisk saks fra mus (tilleggsfigur 1F): Lås inn den distale epifysen mellom bladene og trykk forsiktig mens du snur saksen for å løsne den distale epifysen jevnt uten å kutte opp benet hardt. Hvis det lykkes, skal 4 fremspring være synlige ved den nå eksponerte fysenden (tilleggsfigur 1G). Monter de to lårbenene med den åpne enden vendt nedover i det forberedte 0,5 ml røret plassert inne i et 1,5 ml rør inneholdende FACS-buffer (tilleggsfigur 1H). Plasser en 70 μm cellesil på et 50 ml rør og forvåt silen med 2 ml FACS-bufferen. For å skylle benmargen, sentrifuge rørene (kappene åpne) ved 12.000 x g til sentrifugen når 12.000 x g-verdien, og stopp deretter sentrifugen umiddelbart. Kontroller at benmargscellene er pelletert i 1,5 ml tuben og at lårbenene er hvite (før cellespyling, de er røde) (tilleggsfigur 1I). Kast 0,5 ml rørene med de 2 lårbenene. Kast 150 μl supernatanten med en pipette. Resuspender pelleten med en mikropipette i 1 ml ammoniumklorid-kalium (ACK) lyseringsbuffer i 1-2 minutter ved RT for å lyse røde blodlegemer. Unngå lengre inkubasjonstider, da de kan føre til redusert levedyktighet av kjernefysiske celler. Overfør til 50 ml røret gjennom den forfuktede 70 μm cellesilen. Tilsett 10 ml FACS-buffer for å fortynne ACK-lyseringsbufferen og dermed stoppe lyseringen. Sentrifuge ved 400 x g i 5 minutter ved 4 °C. Resuspender i 10 ml FACS-buffer ved først å resuspendere i 1 ml og deretter fylle på med 9 ml. Forbered cellene for telling som beskrevet i 1.3.8. Tell cellene ved hjelp av en automatisert celleteller etter leverandørens anbefalinger. Det forventes å samle ca 40 millioner celler fra 2 lårben. Farging av benmarg HSPCSentrifuger cellene ved 400 x g i 5 minutter ved 4 °C og resuspender pelleten med en mikropipette i FACS-buffer til en endelig konsentrasjon på 1 x 107 celler/ml. Med en P1000 mikropipette, overfør suspensjonen til et FACS-rør, filtrering gjennom en 35 μm cellesilhette. Forbered enkeltflekkprøverørsprøver for hvert antistoff oppført i tabell 1 for å sette opp kompensasjoner av fluorokromer på cellesortereren:Forbered ett FACS-rør per antistoff og fyll rørene med 200 μL PBS. Tilsett 15 μL fluorokromkompensasjonsperler i hvert FACS-rør av fluorokrom-konjugert antistoff. I FACS-rørene for ufarget og for levende / døde enkeltfargede celler, legg til 500 000 celler i stedet for perler. Tilsett 1 μL av hvert fluorokromkonjugert antistoff (se tabell 1) i det tilsvarende FACS-røret. Tilsett 0,5 μL levende/død flekk i Live/Dead FACS-rør med én flekk. Oppbevares på is i 15 min beskyttet mot lys. Tilbered miks 1 og 2 som angitt i tabell 2.MERK: Antistoffvolumene angitt i tabell 2 er gyldige for antistoffene referert i materialfortegnelsen. De må optimaliseres for enhver ny antistoffreferanse eller et annet parti av samme antistoffreferanse. Tilsett 300 μl Mix 1 i prøverøret, resuspender og oppbevar i 15 minutter på is beskyttet mot lys. Tilsett 300 μl Mix 2 i prøverøret, sett i gang igjen og oppbevar den i 20 minutter på is beskyttet mot lys. Tilsett 3 ml av FACS-bufferen til de enkeltfargede rørene og de blandede prøverørene. Spinn ned ved 400 x g i 5 minutter ved 4 °C. Kast supernatanten forsiktig med en mikropipette og resuspender pelleten i 500 mikrol FACS-bufferen. Forbered et 1,5 ml rør forhåndsfylt med 500 μL oppsamlingsmedium.MERK: Mix 1 er tilberedt i DPBS siden den inneholder Live/Dead-flekken som er betydelig påvirket av HI-FBS. Når cellene er farget av Live / Dead, tilsettes Mix 2, som inneholder fluorokrom-konjugerte antistoffer resuspendert i FACS-buffer som inneholder HI-FBS. Det eneste unntaket er anti-CD16/32 antistoffet som er inkludert i antistoffblanding 1 for å tjene som Fc-reseptorblokker som forhindrer den ikke-spesifikke bindingen av de andre antistoffene som ble lagt til i det følgende trinnet. Cellesortering ved hjelp av en FACSMERK: Mens cellesortering kan utføres på et bredt utvalg av cellesorterere, beskrives her prosedyren for bruk av instrumentene BD FACSAria Fusion eller BD FACSAria III. Det anbefales på det sterkeste at kalibreringen og oppsettet av cellesortereren utføres under tilsyn eller av en erfaren bruker av instrumentet.Kalibrering av FACS-instrument: Se protokoll 1 trinn 4.1. Oppsett av FACS-instrument for cellesortering:Start oppkjøpet av de ufargede cellene. Disse brukes til å definere forover- og sidespredningene og detektorspenningen for hver fluorofor. Sett parametrene slik at det fluorescerende signalet til hver fluorofor faller innenfor det første tiåret av loggskalaen på punktplottet. Skaff enkeltfargekontroller for å definere kompensasjoner manuelt (median for positive og negative populasjoner bør justeres) eller bruk programvaren for automatisk beregning (stigningsmålinger). Kontroller at kompensasjonskontrollene samsvarer med innstillingene for eksperimentelle fluorokrom og detektorer. Registrer 10 000 hendelser for celler og 5 000 hendelser for perler. Bruk prøverøret (dvs. flerfargede celler) for å definere cellepopulasjoner av interesse ved å bruke gating-strategien vist i figur 3A. Følg trinn 4–6 (nedenfor). For å identifisere de tre benmargs-HSPCene av interesse (HSC, CMP og GMP), start gatingen ved å bruke størrelsen (FSC-A) og granulariteten (SSC-A) til gate på leukocytter, deretter FSC-H / FSC-A for å diskriminere dubletter. Basert på SSC-A/dødcellemarkør, gate levende celler. Bruk Lineage/c-Kit til å velge celler som er avstamningsnegative og uttrykker middels til høye nivåer av c-Kit. Gjennom c-Kit / Sca-1, gate på lineage-c-Kit + Sca-1 + (LKS +) HSCs, en av de tre populasjonene av interesse. Blant de myeloide forfedrene (avstamning-c-Kit + Sca-1-), bruk FcγR / CD34 for å ekskludere CD34-FcγR- megakaryocytt og erytroide progenitorer (MEP), mens du inkluderer CD34 + FcγR- CMP, samt CD34 + FcγR + GMP i cellepopulasjonene som skal sorteres. Forsikre deg om at strømmen og nedbøyningen er stabil. I sidestrømkameraet slår du på testsorteringen, spenning PÅ, og bekrefter nøyaktig fallsortering i et 1,5 ml rør montert på venstre side. I vinduet Sorteringsoppsett velger du populasjonen(e) av interesse (dvs. “LKS+” og “CD34+ myeloide progenitorer” vist i dette eksemplet). Under Enhet velger du 2 Tube. Under Presisjon velger du Renhet. I Målhendelser velger du Kontinuerlig for å sortere mellom 160 000 og 200 000 myeloide LKS+ – og CD34+ -stamfedre. Legg til 500 μL FACS-buffer til cellesuspensjonen og overfør 1 ml av prøven ved å filtrere til et nytt 35 μm-cellesilavkortet FACS-rør for å sikre at alle celler er i en enkelt suspensjon like før oppkjøp. Dette eliminerer celleklumper som kan tette instrumentet. Når du er klar, klikker du Sorter og OK for å starte sorteringen. Juster strømningshastigheten for å holde hastigheten under 10 000 hendelser per sekund.MERK: Det forventede forholdet mellom LKS + og CD34 + myeloide progenitorer er 1: 3 for en voksen (8-12 uker gammel) C57BL / 6J kvinnelig mus ved en jevn tilstand. De målrettede sorterte cellenumrene nås vanligvis innen 30 minutter etter sortering. Kvalitetskontroll og telling av sorterte cellerMERK: Dette trinnet skal bare gjøres under piloteksperimentet for optimalisering av prøveprepareringstrinn, med mål om å teste renheten til de sorterte cellene som skal brukes til kjerneisolasjon. Når protokollen er fullstendig optimalisert, anbefales det ikke å utføre dette kvalitetskontrolltrinnet i oppfølgingsforsøkene for å unngå unødvendig sløsing med startmateriale som kan være tilgjengelig i lave antall for kjerneisolering.Renhetskontroll ved flowcytometriOverfør 10 μL av de sorterte cellene til et nytt FACS-rør inneholdende 90 μL FACS-buffer. Innhent og registrer data etter sortering for å bekrefte sorteringsrenhet og levedyktighet. Sørg for at minst 95% av cellene vises i porten av interesse, som definert i 3 – 6 og illustrert i figur 3B. Kjerneisolering fra sorterte benmarg HSPCsBruk protokollen “Low Cell Input Nuclei Isolation” i vedlegget fra 10X Genomics Demonstrated Protocol – Nuclei Isolation for Single Cell Multiome ATAC + GEX sequencing (CG000365 – Rev C)27, med følgende modifikasjoner gjort for å optimalisere kjerneutvinning:Lysistid: Kjør et piloteksperiment for denne protokollen for å identifisere den beste lysetiden for kjerneisolasjon. Sørg for å oppnå en fullstendig cellelyse samtidig som intakte kjerner opprettholdes.MERK: Trinn f i ovennevnte 10X Genomics-protokoll27 instruerer om å “inkubere [i lysisbuffer] i 3-5 minutter på is”. Under pilotforsøket, test minst i 3 minutter, 4 minutter og 5 minutter og vurder gjenvunnet kjernemengde ved telling og kvalitet ved flowcytometri og mikroskopiavbildning for å velge optimal lysvarighet (se beskrivelse av disse kvalitetskontrollene nedenfor). For å spare reagenser, erstatt den fortynnede kjernebufferen med PBS 0,04% BSA i piloteksperimentet. For benmargs-HSPC ble 3 min identifisert som optimal lysisvarighet. Cellesentrifugeringer: For alle celleopphengssentrifugeringer, sentrifuge ved 300 x g i 7 minutter (i stedet for 5 minutter i CG000365 – Rev C)27 ved 4 °C. Kjernesentrifugeringer: Utfør alle kjernesuspensjonssentrifugeringer ved 500 x g i 5 minutter i henhold til CG000365 – Rev C27. Kjerneoppsamling: I trinn b, etter resuspendering i 50 μL PBS 0,04 % BSA og overføring til et 0,2 ml rør, tilsett 50 μL PBS 0,04 % BSA til det originale røret og pipetteblandingen for å samle eventuelle gjenværende celler. Overfør til 0,2 ml rør for å nå totalt 100 μL volum. Heretter vil det totale volumet være 100 μL i stedet for 50 μL i protokollen. Juster trinnene nedstrøms tilsvarende (f.eks. for trinn d, fjern 90 μL i stedet for 45 μL; for trinn e, legg til 90 μL lysisbuffer i stedet for 45 μL). For trinn m, resuspender kjernepelleten i 12 μL fortynnet kjernebuffer i stedet for 7 μL. Tell de isolerte kjernene. I et tomt 0,5 ml rør, tilsett 10 μL 0,4% trypanblå og 8 μL PBS 0,04% BSA. Tilsett 2 μL kjerner til røret og telle kjerner som beskrevet i 1.3.8. Bruk en automatisert celleteller etter leverandørens anbefalinger. Renhetskontroll ved flowcytometriMERK: Dette trinnet skal bare gjøres under piloteksperimentet for optimalisering av prøveprepareringstrinn for å teste renheten til kjernene som skal lastes på 10X Chromium-brikken. Når protokollen er fullstendig optimalisert, anbefales det ikke å utføre dette kvalitetskontrolltrinnet i oppfølgingsforsøkene for å unngå unødvendig sløsing med innsamlede kjerner som kan være tilgjengelige i lave antall.Etter å ha fullført kjerneisolering, overfør 6 μL kjerneresuspensjon i et nytt FACS-rør forhåndsfylt med 150 μL FACS-buffer. Tilsett 3 μL 7-AAD og rug i 5 min på is. Innhent og registrer data etter sortering for å bekrefte sorteringsrenhet og levedyktighet. Sørg for at minst 95 % av kjernene vises i porten av interesse, som definert i protokoll 1 trinn 4.2 (se figur 4). Kvalitetskontroll av rensede kjerner ved mikroskopi:MERK: Dette trinnet skal bare gjøres under piloteksperimentet for optimalisering av prøveprepareringstrinn for å teste kvaliteten på kjernene som skal lastes på 10X Chromium-brikken. Når protokollen er fullstendig optimalisert, anbefales det ikke å utføre dette kvalitetskontrolltrinnet i oppfølgingsforsøkene for å unngå unødvendig sløsing med innsamlede kjerner som kan være tilgjengelige i lave antall.Fortsett som beskrevet i trinn 1.5.3. Utfør multiomanalyseFortsett umiddelbart til Chromium Next GEM Single Cell Multiome ATAC + Gene Expression User Guide (CG000338 – Rev F)25.