단백질-단백질 상호 작용은 면역 조절에서 DNA 복제 및 번역에 이르기까지 대부분의 세포 기능을 강조합니다. 그 결과, 생명과학 전반에 걸쳐 일반적으로 형성되는 다양한 이질적인 복합체에 걸친 광범위한 상호 작용을 조사해야 할 근본적인 필요성이 있습니다. 그러나 이들의 검출, 특성화 및 정량화는 특히 저친화성 상호 작용의 경우 종종 까다롭습니다1.
면역침전 분석은 친화도가 높은 상호작용을 검출하는 데 자주 사용되지만, 친화도가 낮고 일시적인 상호작용의 경우 검출이거의 불가능하다2. 형광 기술도 사용할 수 있지만 형광 라벨을 추가해야 할 수 있습니다2. Cryo-EM은 높은 공간 분해능으로 형성된 단백질 복합체의 구조적 스냅샷과 앙상블 판독을 제공할 수 있지만, 일반적으로 친화성이 낮은 상호 작용을 이미징하기에는 너무 낮은 농도에서 작업해야 합니다. 또한 Cryo-EM은 비용, 접근성, 시료 전처리 및 분석 시간과 관련된 문제를 야기합니다3.
또한 표면 플라즈몬 공명(SPR)은 단백질-단백질 상호 작용을 정량화하는 데 널리 사용되는 방법이 되었지만, 단백질 고정화가 필요하므로 결합 평형에 영향을 미치고 가변 on-rate가 발생하여 측정 정확도가 떨어질 수 있습니다 4,5. 또한 데이터 수집 및 분석 전에 여러 분석 단계가 포함된다6.
질량 측광법은 단백질-단백질 상호 작용을 분석하는 데 사용된 단일 분자 기술입니다 5,6,7. 이는 단일 분자 또는 복합체가 유리 커버슬립(glass coverslip)8의 표면에 닿을 때 산란되는 빛을 기반으로 질량을 측정함으로써 작동한다. 질량 측광 측정은 결합 파트너의 상대적 풍부도와 이들이 형성하는 복합체로부터 결합 친화도를 정량화하는 데 사용되어 왔다5. 그럼에도 불구하고, 다른 단일 분자 기술과 마찬가지로 측정할 샘플의 농도는 일반적으로 100nM 미만이어야 합니다. 농도가 높으면 유리 표면에 착륙하는 분자가 공간적으로 겹쳐져 데이터 품질이 저하됩니다7. 결과적으로, 이러한 낮은 농도에서 해리되는 더 약한 상호 작용(KD ~ 마이크로 몰)은 unbound 종과 bound 종의 필요한 혼합물을 관찰 할 수 없기 때문에 신뢰할 수 있게 측정 할 수 없습니다5.
여기에서는 새로운 결합 된 미세 유체 질량 측광 장치를 기반으로 이러한 한계를 극복하는 접근 방식을 설명합니다. 특히, 미세유체역학 시스템은 질량 광도계와 함께 사용되어 질량 측광으로 정량화할 수 있는 상호 작용의 범위를 효과적으로 확장합니다. Microfluidics는 약한 상호 작용 1,9를 검출하기 위하여 급속한 희석을 포함하여 단백질 단백질 상호 작용 조사를 조사하기를 위한 가능성의 범위를 제안하기 위하여 보였습니다. 본원에 기술된 시스템은 미세유체 칩에서 샘플을 최대 10,000배까지 빠르게 희석하고 칩의 관찰 영역을 가로질러 즉시 흐르게 함으로써 작동하며, 분자가 희석 과정(10)을 시작한 시점으로부터 50ms 이내에 질량 측광 측정을 시작할 수 있도록 한다. 희석은 샘플과 완충액이 칩의 역 Tesla 밸브 믹서에서 결합될 때 발생하며, 두 용액의 상대 유속에 따라 발생하는 희석량이 결정됩니다(프로토콜 8단계 참조). 유속은 미세유체 제어 소프트웨어로 제어할 수 있습니다. 유속을 변경하면 질량 광도계로 측정되는 유리 표면의 착륙 이벤트 수에 영향을 줄 수 있으므로 종의 상대적 개체수가 변경될 수 있습니다.
프로세스의 속도는 상호 작용의 무결성이 중단되기 전에 측정을 완료할 수 있을 만큼 충분히 빠릅니다(자세한 내용은 토론 참조). 이것은 1차 반응 이론을 간략하게 살펴보면 이해할 수 있습니다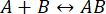 . 순방향(연관성) 속도 상수는 kf이고, 역방향(해리) 속도 상수는 kb이며, 평형 해리 상수(KD)는 다음과 같이 정의됩니다.
. 순방향(연관성) 속도 상수는 kf이고, 역방향(해리) 속도 상수는 kb이며, 평형 해리 상수(KD)는 다음과 같이 정의됩니다.
케이D= kb/ kf
단백질 결합의 경우, kf는 일반적으로 반응물의 확산(11)에 의해 제한되며, 따라서 10,6-10,7M-1·s-1의 범위로 제한된다. 의 범위가 제한되어 있기 때문에 친화성이 낮은(KD~마이크로몰) 반응은 kb≈ 1 s-1을 갖습니다. 즉, kb= kf · KD = (106 M-1·s-1) (10-6 M) = 1 s-1, 복합체의 반감기는 약 0.7 s11,12입니다.
본 예시 시스템은 IgG 단클론 항체 트라스투주맙(trastuzumab)을 상호작용 파트너로 알려진 IgG 신생아 Fc 수용체(FcRn)의 용해성 도메인에 결합하는 것이다13. 기존의 질량 측광법을 단독으로(즉, 샘플의 수동 희석으로) 얻은 이전에 발표된 데이터는 단백질이 여러 종을 형성한다는 것을 보여주었습니다. FcRn 단량체, FcRn 이량체 및 결합되지 않은 IgG를 명확하게 볼 수 있었으며, IgG-FcRn 복합체(1:1 및 1:2 비율)도 검출되었지만(pH 5.0에서) 매우 낮은 농도에서만 검출되었습니다5. 이 관찰은 더 높은 농도에서 측정할 경우 IgG-FcRn 복합체 형성이 더 명확하게 검출될 수 있는지에 대한 질문을 제기합니다. 실제로, 여기에 설명된 질량 측광법과 결합된 급속 희석 접근법을 결합하면 측정된 입자의 증가에 의한 복잡한 형성에 대한 보다 강력한 증거를 제공할 수 있습니다.
여기에 설명된 질량 측광 및 미세유체역학 프로토콜은 마이크로몰 범위까지 KD 를 가진 복합체의 형성을 특성화하는 것을 가능하게 합니다. KD 의 경험적 측정은 유량 센서 정확도, 펌프 안정성, 칩 간 변동 및 관찰 창 내의 측정 위치에 대한 추가 개선이 필요하며, 이러한 모든 요인이 샘플이 희석된 시점부터 측정까지의 시간에 영향을 미치기 때문입니다.
질량 광도계(30 kDa – 6 MDa)로 분석하기에 적합한 범위에 속하는 고유한 분자량(최소 25kDa로 분리)이 있는 경우 용해성 단백질 간의 결합을 조사하기 위해 동일한 접근 방식을 적용할 수 있습니다. 얻은 통찰력은 세포 기능에 대한 기계론적 이해를 얻는 것부터 새로운 바이오 치료제 설계에 이르기까지 다양한 맥락의 연구에 도움이 될 수 있습니다.
여기에 설명된 프로토콜은 저친화성 단백질-단백질 상호 작용을 검출하고 정량화하는 방법을 제공합니다. 급속 희석 미세유체 시스템에 결합된 질량 광도계를 사용합니다. 질량 측광법은 30 kDa에서 6 MDa 범위 내의 생체 분자16에 대한 용액에서 분자 질량을 신뢰성 있게 측정할 수 있는 비표지 생물 분석 도구입니다. 질량 측광법은 샘플을 하나씩 분석하는 단일 분자 기술이므로 일반적으로 100pM-100nM 농도 범위의 샘플로 제한됩니다. 이 범위를 벗어나면 유리 표면에 착륙하는 분자가 공간적으로 겹쳐져 데이터 품질이 저하됩니다. 이 범위 이하에서는 얻은 데이터가 너무 적어 강력한 분석을 수행할 수 없다7. 중요한 결과는 단백질 상호 작용에 대한 연구를 해당 범위 내에서 결합 된 종과 결합되지 않은 종의 혼합물을 형성하는 것으로 제한 할 수 있다는 것입니다.
여기에서는 대량 광도계에 적합한 시료 농도 범위를 효과적으로 확장하기 위해 급속 희석 미세유체 시스템을 사용하기 위한 단계별 프로토콜에 대해 자세히 설명했습니다. 미세유체 칩에서 시료를 희석한 다음 50ms 이내에 검출기 관찰 창을 가로질러 흐르게 함으로써 시스템은 상호 작용 평형이 이동하기 전에 희석되지 않은 시료에 존재하는 복합체를 캡처합니다. 샘플은 개별 측정 중에 지속적으로 검출기로 전달됩니다. 이러한 조건에서 복합체의 95%는 샘플을 측정할 때 손상되지 않은 상태로 유지되며, 낮은 친화성 상호 작용의 경우에도 미세몰 순서에 따라 KD 가 있고 해리 속도가 1 s-1만큼 빠릅니다.
이는 다음과 같이 계산할 수 있습니다: 순방향 속도 kf및 역방향 속도 kb를 갖는 반응 에 대해,
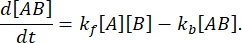
평형 상태에서는 세 종(A, B 및 복합체 AB)의 농도가 모두 일정하게 유지되므로 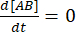 및
및 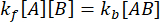 . 섭동(이 경우 희석)으로 인해 복합체가 해리될 수 있지만 순방향(결합) 반응이 진행되지 않는다는 보수적 가정 하에서 kf [A] [B]라는 항은 무시할 수 있는 것으로 취급될 수 있으며 다음과 같이 단순화할 수 있습니다.
. 섭동(이 경우 희석)으로 인해 복합체가 해리될 수 있지만 순방향(결합) 반응이 진행되지 않는다는 보수적 가정 하에서 kf [A] [B]라는 항은 무시할 수 있는 것으로 취급될 수 있으며 다음과 같이 단순화할 수 있습니다.
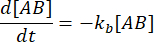
적분은 평형의 섭동 이후의 시간에 복합체의 농도에 대해 다음과 같은 표현을 제공합니다.
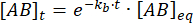
평형의 섭동 후 시간 t 에 묶인 상태로 유지되는 복합체의 비율은 다음과 같습니다.
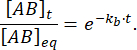
= 50ms에서 kb≈ 1 s-1과의 반응에 대해 분율 한계는 0.95 또는 95%11,12입니다.
질량 측광법은 FcRn의 용해성 도메인에 대한 IgG 단클론 항체 trastuzumab의 결합을 조사하기 위해 여기와 이전에5 를 사용했습니다. 2개의 의무적인 파트너는 산성 PH17에 nanomolar 친화도로 결합하기 위하여 보고되었습니다. 질량 측광법은 결합 파트너가 pH 5.0에 있을 때 형성된 복합체의 풍부도를 정성적으로 평가하는 데 사용되었으며, 샘플은 추가 미세유체 시스템을 통해 빠르게 희석되었습니다. 이 절차는 이전에 보고된 결과를 기반으로 특정 단백질-단백질 상호 작용에 최적화되었습니다5. 사용자가 사전 지식이 있거나 사용할 완충액, 초기 단백질 농도, 예상 화학량론, 상호 작용이 평형에 도달하는 데 필요한 배양량과 같은 해당 시스템의 실험 조건을 최적화하는 경우 동일한 절차를 사용하여 다른 상호 작용을 연구할 수 있습니다.
IgG-FcRn 혼합물을 수동으로 희석했을 때, 이러한 단백질이 상호 작용하는 것으로 알려져 있음에도 불구하고 IgG-FcRn 복합체의 존재를 감지하기 어려웠습니다5. 이 논문은 급속 희석 접근법이 이러한 복합체의 양을 현저히 증가시킨다는 것을 보여줍니다. 동일한 샘플에 대해 급속 희석을 사용했을 때 1:1 FcRn-IgG 복합체와 2:1 FcRn-IgG 복합체가 모두 명확하게 관찰되었습니다. 복잡한 형성의 이러한 차이는 광범위한 농도에 걸쳐 생체 분자 상호 작용 시스템을 연구하는 것의 중요성을 보여줍니다.
게다가, 이러한 결과는 또한 단일 분자 분석과 함께 미세유체역학을 사용하여 약한 상호 작용을 포착하는 것이 간단하다는 것을 보여줍니다. – 방법의 상당한 격차를 메우는 것. 급속 희석 미세유체역학과 질량 측광법의 조합은 분석 기술로서 질량 측광법의 장점으로 인해 매력적인 이점을 제공합니다. 즉, 질량 측광은 라벨이 필요하지 않고 최소한의 시료 준비만 필요하며 측정은 용액에서 수행됩니다. 이 프로토콜의 경우 질량 측광의 또 다른 주요 이점은 형성된 모든 종을 구별하고 정량화할 수 있다는 것입니다(>30kDa의 고유한 질량을 갖는 경우). 예를 들어, SPR은 결합 및 결합 해제 속도를 측정할 수 있지만 화학량론 정보를 쉽게 제공할 수 없는 것과는 대조적이다8.
이 프로토콜과 보다 일반적으로 대량 측광 실험의 경우 몇 가지 고려 사항이 도움이 됩니다. 첫째, 최종 단백질 농도는 질량 측광법이 측정할 수 있는 한계(100 pM-100 nM) 내에 있어야 합니다. 시작 배양 농도는 또한 미세유체 시스템의 범위(최대 90μM) 내에 있어야 하며 상호 작용10의 실제 KD보다 높은 것으로 이론화됩니다. 권장되는 시작점은 μM 농도에서 상호 작용하는 종 간의 1:1 농도 혼합 비율입니다. 그런 다음 비율을 1:2, 1:5 또는 이 상호 작용의 경우와 같이 1:10으로 변경할 수 있습니다. 단백질 상호 작용에 대한 이전 정보가 없는 경우 사용자는 구성 요소의 친화도가 제시된 방법에 의해 유지되는 농도 범위 내에 있는지 확인(즉, 복합체가 형성됨)하기 위해 각 파트너에 대해 고농도(권장 20μM)부터 시작하여 실험을 최적화해야 합니다. 최적화에는 올바른 혼합 비율을 결정하기 위해 상호 작용 성분 중 하나의 상호 작용 또는 적정을 촉진하기 위해 다른 완충액 조건을 선택하는 것도 포함될 수 있습니다. 이러한 사항이 결정되면 농도와 흐름을 최적화하여 연구 및 방법에 대한 최적의 조건을 허용할 수 있습니다(예: 더 나은 피크 분리능을 허용하기 위해 농도를 줄이는 것).
둘째, 이 실험을 성공적으로 재현하려면 불순물을 최소화해야 합니다. 질량 측광 측정에 부정적인 영향을 미치는 것으로 알려진 불순물의 일반적인 원인에는 정제 후 남아 있는 다른 단백질 또는 세포 파편, 여과되지 않은 완충액, 미셀 형성 세제(너무 높은 농도로 존재하는 경우) 및 고농도의 염, 글리세롤 또는 기타 성분을 포함하는 완충액이 포함됩니다. 위의 프로토콜에서 논의된 바와 같이 미세유체역학 시스템의 기포를 제거해야 합니다. 튜브 시스템에 기포가 형성되거나 샘플의 표면 장력이 높고 거품이 형성되기 쉬운 경우 기포가 형성될 수 있습니다. 이멀젼 오일에도 기포가 형성될 수 있으며, 이는 초점 링에서 감지할 수 있습니다(그림 3). 프로토콜에 설명된 단계를 사용하여 기포를 제거할 수 없는 경우 또 다른 해결책은 데시케이터와 진공 펌프를 사용하여 샘플의 가스를 제거하고 샘플을 몇 분 동안 감압 상태로 두는 것입니다. 고농축 단백질 용액을 소용돌이치거나 흔드는 것은 기포 형성을 촉진할 수 있으므로 권장되지 않습니다.
하나의 특정 단백질-단백질 상호 작용에 대한 측정이 여기에서 입증되었지만, 동일한 프로토콜을 큰 수정 없이 다른 단백질-단백질 상호 작용 시스템에 적용할 수 있습니다. 이 프로토콜의 또 다른 미래 방향은 질량 측광 5,7의 맥락에서 다른 곳에서 설명된 바와 같이 식별된 복합체에대한 KD 값을 계산하기 위해 측정값을 사용하는 것입니다. 이전 연구에서는 수동 희석 및 더 강력한 상호 작용과 관련된 실험 데이터를 사용했지만, 미세유체 장치의 추가 개선이 구현되는 경우(예: 유량 센서 정확도 및 펌프 안정성 향상) 분석 원리를 이러한 맥락에서 쉽게 적용할 수 있습니다.
단백질-단백질 상호 작용 외에도 결합 질량 측광 및 급속 희석 미세유체역학 접근 방식에 대한 더 넓은 응용 분야가 있을 수 있습니다. 질량 측광법은 시료 순도, 응집 및 균질성을 평가하는 데 사용할 수 있습니다18,19; 단백질 올리고머화20, 고분자 조립21 또는 중합22를 연구합니다. 그리고 다른 지역에서. 질량 측광 분석은 단백질에만 국한되지 않습니다. 핵산과 단백질23, 바이러스 입자24 및 나노 입자25 사이의 상호 작용을 조사하는 데 사용되었습니다. 따라서 이 프로토콜은 결합 질량 측광 미세유체역학 시스템의 중요한 응용 분야를 설명합니다 – 이를 통해 개별 분자 및 복합체 수준에서 약한 단백질-단백질 상호 작용을 직접 측정할 수 있습니다. 본 응용 프로그램의 가치는 일반적으로 연구하기 어려웠던 상호 작용을 직접 특성화할 수 있는 가능성을 열어주기 때문에 매우 높으며, 이는 중요한 치료 영역 전반에 걸쳐 관련성이 있습니다. 이 결합된 접근 방식은 최대 수십 마이크로몰 농도의 샘플에 대한 광범위한 조사의 기초가 될 수도 있습니다.
