Figure 4A shows spontaneous spikes that arise from a sensillum. They fall into two classes based on amplitude, with the larger spikes deriving from the neuron that is sensitive to bitter compounds and the smaller spikes from the neuron that responds to sugars. The relationship between spike amplitude and functional specificity has been corroborated by genetic experiments4,14,37,38,39 .
Figure 4B shows the response of the bitter-sensitive neuron of the S5 sensillum to the odor of DEET; this response occurred without any contact between the DEET solution and the sensillum. One spike appears to be of intermediate amplitude and may result from the superposition of two smaller spikes, one from the sugar neuron and one from the mechanosensory neuron.
Figure 4C shows the ON response that occurs following contact between a bitter stimulus and the I1 sensillum, followed by an OFF response that occurs following the termination of contact. Not that the magnitude of the OFF response, measured in spikes/s, is greater than the ON response.
Figure 4D shows an ON and an OFF response to the bitter compound berberine from an I1 sensillum of another species of fly, Drosophila virilis. One of the advantages of this technique is that it can be performed on other species, including mosquitoes, without the need to introduce any transgenes into them.
Figure 4E illustrates two problems that can occur during recording. First, if the labellum is not properly secured it can move, producing spikes from the mechanosensory neuron of the sensillum. Second, the recording electrode can become dislodged from the sensillum, often as a result of movement of the labellum. In this case, contact is lost and the electrode must be re-inserted to record spikes.
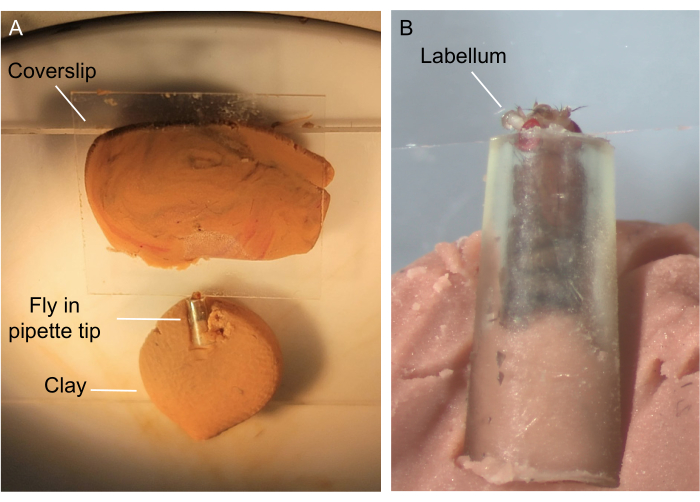
Figure 1: Fly preparation for base recording. A trimmed pipette tip containing a female fly with the labellum securely placed on a glass coverslip. (A) Both the pipette tip and the coverslip are held securely on mounds of clay. (B) Higher magnification of labellum resting on a coverslip. Please click here to view a larger version of this figure.

Figure 2: Electrophysiology rig setup. (A) An overview showing the positions of a manual micromanipulator for the reference electrode, a motorized micromanipulator for the glass stimulus capillary, and a motorized micromanipulator for the recording electrode. (B) Overview showing holders for the reference electrode, glass stimulus capillary, and recording electrode. Please click here to view a larger version of this figure.
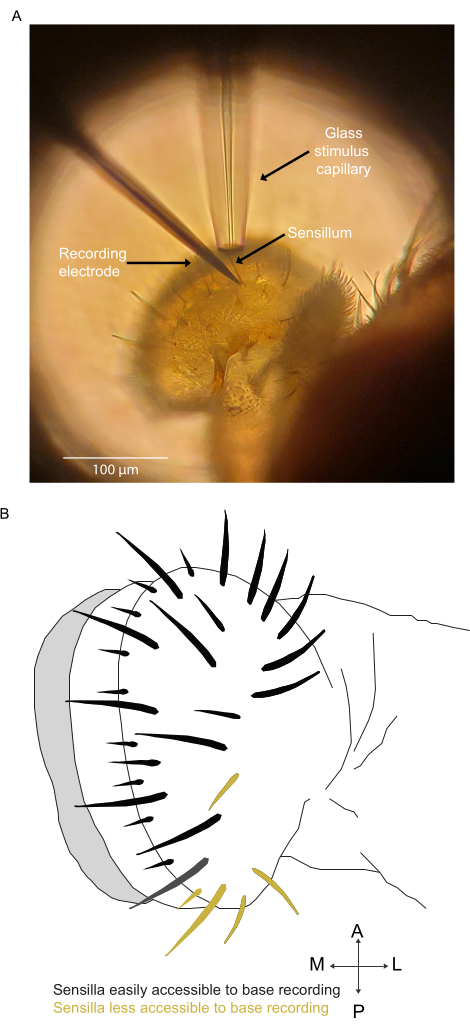
Figure 3: The labellum and sensilla. (A) The labellum under an inverted microscope. Close-up of the labellum showing the taste sensilla of one lobe of the labellum. It also shows the stimulus glass capillary close to one of the large sensilla known as L2 (Large type 2) and a recording electrode in the base of this sensillum. (B) Accessibility of labellar sensilla. The labellum showing sensilla that are easily accessible to base recording and sensilla that are less accessible due to their position in the preparation that we generally use. Abbreviations: A = anterior; M = medial; P = posterior; L = lateral. Please click here to view a larger version of this figure.
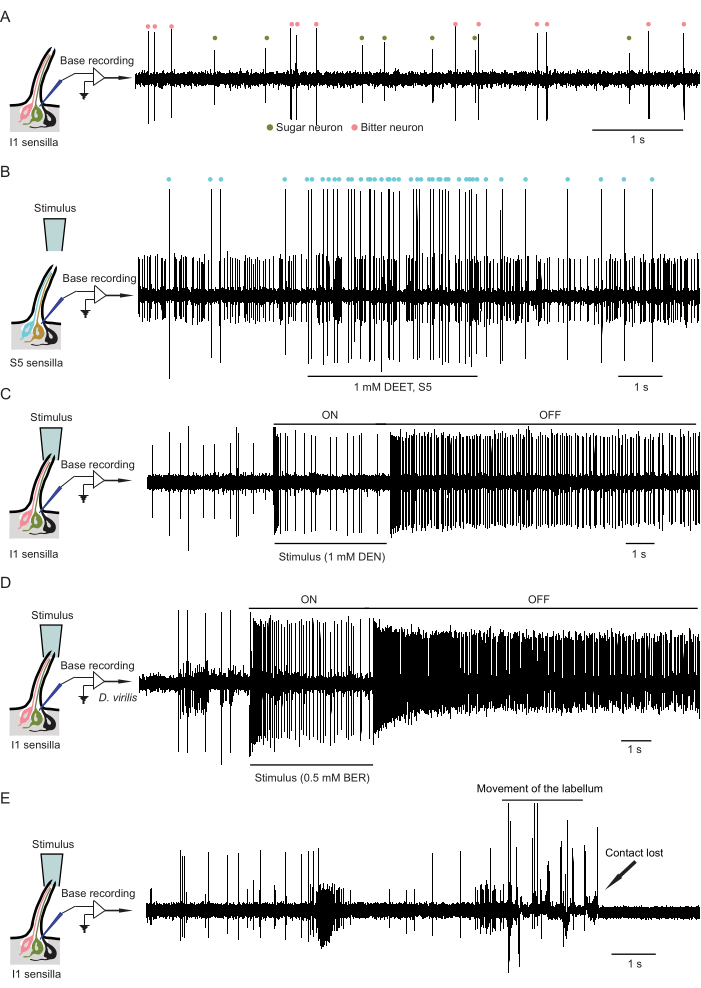
Figure 4: Example traces of spontaneous spikes, bitter neuron responses, and suboptimal recording. (A) Example trace of spontaneous spikes. The spikes are from a bitter-sensitive neuron (red dots) and a sugar-sensitive neuron (green dots) in an I1 sensillum. In the drawing of the sensillum, the bitter, sugar, and mechanosensory neurons are colored red, green, and black, respectively. (B) Example trace of the response of the bitter neuron (red dots) in an S5 sensillum to the vapor of 1 mM DEET. Note that the small amplitude spikes are from the sugar neuron and mechanosensory neurons, which in this example trace are difficult to distinguish by amplitude. Abbreviation: DEET = N,N-diethyl-meta-toluamide. (C) Example trace of ON and OFF responses from a bitter neuron in I1 sensilla to 1 mM denatonium benzoate. Spikes are observed before (spontaneous firing), during (ON response), and after (OFF response) contact with the stimulus. Abbreviation: DEN = denatonium benzoate. (D) Example trace of ON and OFF responses from a bitter neuron in I1 sensilla of D. virilis to 0.5 mM berberine chloride. Spikes are observed before, during, and after contact. Abbreviation: BER = berberine chloride. (E) Example trace of a suboptimal recording. The contact was lost in the middle of the experiment due to the movement of the labellum. Please click here to view a larger version of this figure.
