1. Genomic DNA Fragmentation
Fragment genomic DNA using sonication to a desired size range suited for the genome-wide sequencing platform. (We usually sonicate to ~300 bp.) Verify the size distribution of the fragmented genomic DNA on 1% agarose gel (Figure 1).
2. DNA Preparation
Determine the starting DNA amounts based on the abundance of 5-hmC in genomic DNA. Since 5-hmC levels vary significantly in different tissue types, starting DNA amounts depend on the 5-hmC levels of the samples. Please refer to Table 1 for examples.
3. β-GT Catalyzed Reaction (Glucose Transfer Reaction)
- Mix by pipetting the mixture as detailed in Table 2 and incubate in a 37 °C water bath for 1 hr.
- After incubation, clean up the reaction with QIAquick Nucleotide Removal Kit, using 10 μg of DNA per column. Elute with 30 μl water per column and combine.
4. Biotinylation Reaction (Click Chemistry)
- Add DBCO-S-S-PEG3-Biotin conjugate working solution (1 mM) in the eluted DNA solution (from step 3) to a final concentration of 150 μM (i.e., 5 μl of working solution per 30 μl DNA solution).
- Mix by pipetting and incubate in a 37 °C water bath for 2 hr.
- Clean up the reaction with QIAquick Nucleotide Removal Kit. The ideal total elution volume is 100 μl.
- Quantify the recovered DNA amount using microliter scale spectrophotometer (e.g., NanoDrop).
5. Capture of 5-hmC-containing DNA
- Wash 50 μl of Dynabeads MyOne Streptavidin C1 3 times with 1 ml of 1X B&W buffer according to the manufacturer’s instructions. Separate the beads with a magnetic stand.
- Add equal volume of 2X B&W buffer to the recovered biotinylated DNA (100 ul) to the washed beads.
- Incubate for 15 min at room temperature with gentle rotation on a rotator.
- Separate the beads with a magnetic stand and wash the beads 3 times with 1 ml of 1X B&W buffer.
- Elute the DNA by incubating the beads in 100 μl of freshly prepared 50 mM DTT for 2 hr at room temperature with gentle rotation on a rotator.
- Separate the beads with a magnetic stand. Aspirate the eluent and load onto a Micro Bio-Spin 6 Column according to the manufacture instruction to remove the DTT. The target DNA is in the solution now.
- Purify the eluted DNA from the previous step by Qiagen MinElute PCR Purification Kit and elute DNA in 10 μl of EB buffer. Quantify DNA using Qubit Fluorometer, or NanoDrop if concentration is higher than 20 ng/ul. The DNA is ready for downstream genome-wide sequencing library preparation.
6. Representative Results
If the quality of genomic DNA is high, typical recovery yields after the β-GT and biotinylation reactions are ~60-70%. However, the capture efficiency vary significantly with different tissue types depending on the 5-hmC levels of the samples. Typically, the capture efficiency for brain genomic DNA is ~4-9%, and in some extreme cases, the efficiency may reach up to 12%. For ES cells, the average capture efficiency is ~2-4%, in contrast to ~0.5% for neural stem cells. The lowest efficiency seen so far was for genomic DNA from cancer cells. All enriched DNA is ready for standard next-generation library preparation protocols. In addition, the captured DNA can also be used as template for real-time PCR to detect the enrichment of some fragments compared to the input DNA, if the related primers are available.
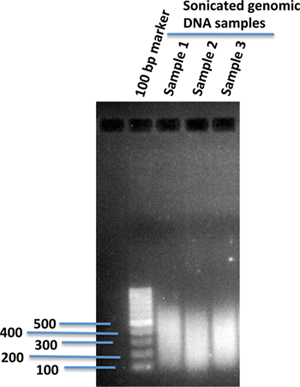
Figure 1. Sonicated human genomic DNA fragments in 1% agarose gel. 10 μg of genomic DNA isolated from human iPS cells in 120 μl of 1X TE buffer was sonicated using a sonication device (Covaris). After sonication, 2 μl of the sonicated DNA was loaded onto 1% agarose gel using 100 bp of DNA marker to compare the sizes of the sonicated DNA fragments.
| Component | Volume | Final Concentration |
| Water | _ μl | |
| 10 X β-GT Reaction Buffer | 2 μl | 1 X |
| Up to 10 μg genomic DNA | _ μl | Up to 500 ng/μl |
| UDP-6-N3-Glc (3 mM) | 0.67 μl | 100 μM |
| β-GT (40 μM) | 1 μl | 2 μM |
| Total volume | 20 μl |
i) For tissue genomic DNA (high 5-hmC content > 0.1%)
| Component | Volume | Final Concentration |
| Water | _ μl | |
| 10 X β-GT Reaction Buffer | 10 μl | 1 X |
| Up to 20 μg genomic DNA | _ μl | Up to 500 ng/μl |
| UDP-6-N3-Glc (3 mM) | 1.33 μl | 100 μM |
| β-GT (40 μM) | 2 μl | 2 μM |
| Total volume | 40 μl |
ii) For stem cell genomic DNA (median 5-hmC content ~0.05%)
| Component | Volume | Final Concentration |
| Water | _ μl | |
| 10 X β-GT Reaction Buffer | 10 μl | 1 X |
| Up to 50 μg genomic DNA | _ μl | Up to 500 ng/μl |
| UDP-6-N3-Glc (3 mM) | 3.33 μl | 100 μM |
| β-GT (40 μM) | 5 μl | 2 μM |
| Total volume | 100 μl |
iii) For cancer cell genomic DNA (low 5-hmC content ~0.01%)
Table 1. Examples of amounts of input DNA and labeling reactions using the samples with various 5-hmC levels by the selective chemical labeling method.
| Sample | 5-hmC level | Starting DNA (μg) | Recovery after labeling (input to beads) (μg) | Recovery yield | Pull-down DNA (ng) | Pull-down yield |
| Adult mouse cerebellum | 0.4% | 10 | 7.5 | 75% | 236 | 3.1% |
| Postnatal day 7 mouse cerebellum | 0.1% | 11 | 9 | 82% | 140 | 1.6% |
| Mouse ES cell E14 | 0.05% | 60 | 42 | 70% | 350 | 0.8% |
Table 2. Representative results from mouse brain tissues and ES cells.