Typical results show βAPP leaving the TGN and appears to traffic rapidly to LAMP1 (Figure 1a and b). During the photo-activation period, vesicles can be seen departing the Golgi destined for lysosomes (Figure 1b). Without inhibitor treatment, paGFP fluorescence will be visible in lysosomes while there is photo-activation in the TGN. After stopping photoactivation, the βAPP-paGFP is rapidly cleared from the lysosome (compare Figure 1a to 1b). Treatment with nocodazole leads to the accumulation of APP within the GalT-CFP labeled compartments and prevents trafficking to lysosomes (Figure 1c).
The Swedish mutation of APP (APPsw) increases the rate of β-cleavage by 10-fold 34. With rapid cleavage of βAPPsw-paGFP, the paGFP fluorescence appears as a diffuse fluorescence signal in the cytosol. Presumably this is the result of rapid βAPP cleavage by γ-secretase liberating the C-terminal paGFP in to the cytoplasm (Figure 2). In the presence of γ-secretase inhibitor, βAPP accumulates within lysosomes.
After delivery of βAPP to the lysosome, the paGFP fluorescence is cleared rapidly. The short residence time could be the result of trafficking from the lysosome to another compartment. Conversely, cleavage of βAPP by γ-secretase would be expected to lead to diffusion of paGFP fluorescence from the lysosomal membrane (Figure 3a). The diffuse fluorescence is more difficult to detect by confocal microscopy. To determine whether βAPP is delivered to another compartment or cleared, SN56 cells were treated with a specific inhibitor against γ-secretase (L685, 458) or an alkalinizing agent (chloroquine). The addition of either L685, 458 or chloroquine resulted in APP accumulation in the lysosome (Figure 3b and c).
It is commonly accepted that APP is delivered to the cell-surface before being endocytosed and processed in endosomes and lysosomes 35. However, in our untreated cells, cell-surface βAPP could not be detected by confocal microscopy. Photo-activated βAPP may not be concentrated in a specific region of plasma membrane. Instead, there may be a diffuse fluorescence across the large surface area of the plasma membrane. Interestingly, γ-secretase inhibitor treatment leads to detectable βAPP-paGFP at the cell surface (Figure 3b). γ-secretase inhibitor treatment may route more βAPP to the cell surface, in addition to inhibiting enzyme function. Therefore, the large surface area of the plasma membrane spreads paGFP over a larger area and makes detection difficult, although the protein is trafficked to the compartment.
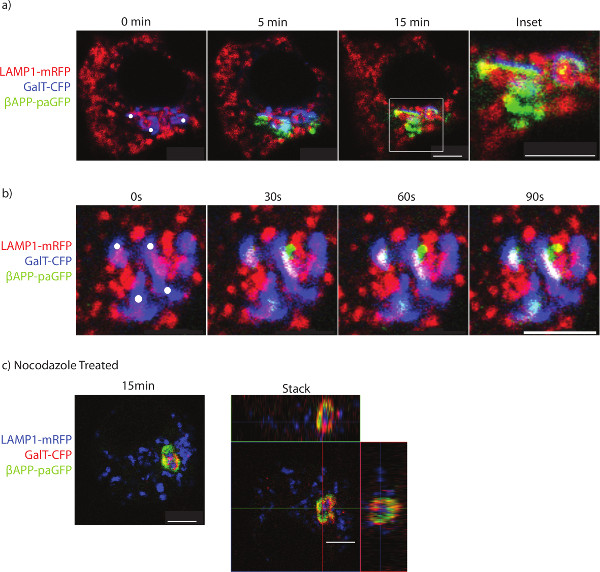
Figure 1. Imaging the trafficking of APP to lysosomes. SN56 cells were transfected with LAMP1-mRFP, GalT-CFP, and βAPP-paGFP. a) The cell was accurately photo-activated within ROIs placed over the Golgi (ROIs denoted by white circles). The cell was alternatively photo-activated and imaged for 15 min to determine βAPP-paGFP trafficking. Inset shows trafficking within the perinuclear area from Golgi to lysosomes. Co-localized pixels appear yellow. b) The trafficking of a vesicle containing βAPP-paGFP to the lysosome. Yellow pixels denote LAMP1 and βAPP-paGFP colocalization. c) SN56 cells were treated with nocodazole before imaging to prevent βAPP-paGFP egress from the TGN. A representative image taken from the end of the photo-activation period. In the right panel, a Z-stack of the same cell taken immediately after photo-activation. The GalT-CFP has been false colored red to improve visualization of colocalized GalT-CFP and βAPP-paGFP. Yellow pixels denote colocalization between GalT-CFP and βAPP-paGFP. Scale bars represent 5 µm. Please click here to view a larger version of this figure.
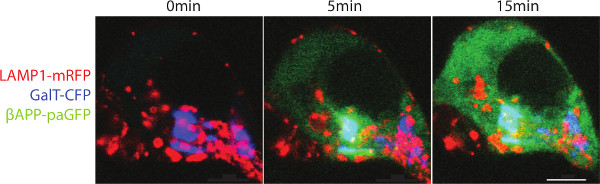
Figure 2. Trafficking of βAPPsw-paGFP. SN56 cells were transfected with LAMP1-mRFP, GalT-CFp and βAPPsw-paGFP (βAPP with the familial Swedish mutation). The Swedish mutation increases the rate that APP is cleaved. Cells were alternatively within the Golgi photo-activated and imaged. A diffuse fluorescence can be seen after paGFP is detached from the lysosomal membrane to diffuse into the cytoplasm. Scale bar represents 5 µm. Please click here to view a larger version of this figure.
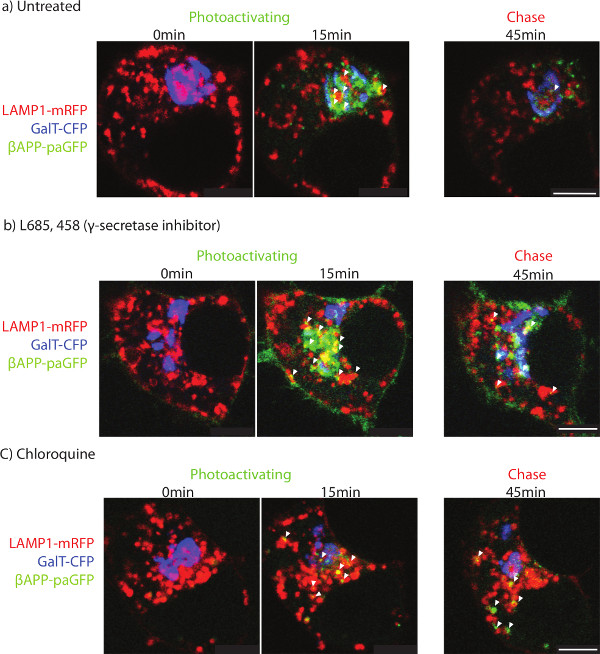
Figure 3. Determining the terminal organelle. SN56 cells were transfected with LAMP1-mRFP, GalT-CFP and βAPP-paGFP. After photo-activation, cells were followed for an additional 45 min. The two left-most panels show the cells before photoactivation (left-most) and 15 min after photoactivation (middle). The panel on the far right shows APP fluorescence after 45 min of total imaging in a) untreated, b) γ-secretase inhibitor treated, or c) chloroquine treated. Arrowheads point to APP co-localized with LAMP1 after 45 min of imaging. Scale bars represent 5 µm. Please click here to view a larger version of this figure.