Sub-mitochondrial localization of CyclinB1 and Cdk1
Sodium carbonate extraction is used to determine whether a protein is located inside the mitochondria or on the outside surface, namely outer membrane. Once a protein is shown to localize inside the mitochondria, further determination of sub-mitochondrial localization can be made via mitoplasting combined with protease digestion. To specify the sub-mitochondrial localization of CyclinB1 or Cdk1, mitoplasts were isolated by diluting mitochondria in hypotonic buffers with decreasing concentrations of the osmotic sucrose from 200 mM to 25 mM. The outer membrane begins to rupture at 150 mM of sucrose, while the inner membrane remains intact until the final concentration at 25 mM of sucrose (Figure 1A). In combination with mitoplasting, protease protection assay can be performed using trypsin to digest exposed proteins following outer membrane rupture. This will result in digestion of intermembrane space proteins. If the protein of interest is protected from trypsin digestion, this indicates mitochondrial matrix localization of the protein. In this representative figure, mitochondrial matrix protein Hsp60, and intermembrane space protein Timm13 were used as sub-mitochondrial localization markers. Similar with Hsp60 but unlike Timm13, CyclinB1 and Cdk1 were protected from trypsin digestion, indicating their mitochondrial matrix localization (Figure 1B).
Mitochondrial Expression of MTS- and GFP-tagged CyclinB1 and Cdk1 Proteins
MTS is cloned in frame at the N-terminus of CyclinB1 or Cdk1 genes, which has GFP or RFP tags at their C-terminus. The resultant recombinant protein is mitochondria-targeted GFP- or RFP-tagged CyclinB1 or Cdk1. The list of the constructs generated and used in this study is shown in the figure. Using these constructs, overexpression of CyclinB1 and/or Cdk1 in the mitochondria was achieved, shown here by western blotting of the isolated mitochondrial fractions (Figure 2).
Potential Mitochondrial Targets of CyclinB1/Cdk1 Determined by 2D-DIGE
Cdk1 belongs to the serine/threonine (S/T) kinase family catalyzing the transfer of a phosphate from ATP to proline (P)-oriented S or T residues. A point mutation that replaces an aspartate (D) residue with asparagine (N) at position 146 of Cdk1 (D146N) generates a dominant negative (dn) Cdk1 mutant19. To study the function of mitochondrial Cdk1, a mitochondria-targeted Cdk1-dn protein was generated by constructing a plasmid (pERFP-N1-MTS-Cdk1-dn) containing a 29 amino acid-long mitochondrial targeting sequence (MTS) derived from the subunit VIII of the human cytochrome C oxidase linked to RFP-tagged dn-Cdk1. pERFP-N1-MTS producing mitochondria-targeted ERFP protein was used as an empty vector control. Mitochondrial phosphoproteins in G2/M cells transfected with both constructs were profiled by 2D gel analysis with pH 4-10 gel strips. Compared with empty vector transfectants (Figure 3, upper panel), a group of mitochondrial phosphoproteins was apparently absent or decreased in the Cdk1-dn transfectants (Figure 3, lower panel). Mass spectrometry analysis of the spots detected determined the identity of the proteins phosphorylated by Cdk1 in the mitochondria.
Cell Cycle Progression and Determination of Phase Lengths with EdU Pulse-chase Assay
To investigate the progression of cell cycle when mitochondrial CyclinB1/Cdk1 levels are increased, a pulse-chase labeling experiment using a thymidine analogue, ethynyl deoxyuridine (EdU) was performed to label the population of cells undergoing DNA synthesis20. This method allows the visualization of cell cycle captured over a 22 hr window by tracking the EdU-positive population when cells progress through S and G2/M phases and accumulate in G1 phase. The results show that labeled S phase cells progressed through G2/M phase and appeared in G1 phase as fast as 4 h in cells expressing wild type mitochondrial CyclinB1/Cdk1, as compared to 6 h in cells transfected with a vector control or mutant CyclinB1/Cdk1 (Figure 4A), indicating that enhancement of mitochondrial CyclinB1/Cdk1 accelerates cell cycle progression.
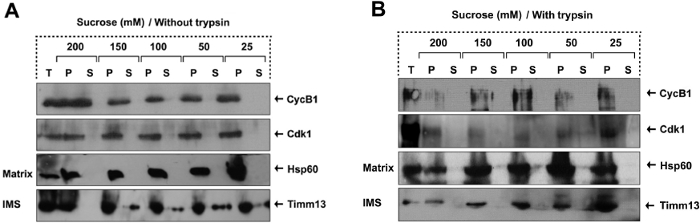
Figure 1. Mitochondrial CyclinB1/Cdk1 Localizes in the Matrix. (A-B) Sub-mitochondrial localization of CyclinB1 and Cdk1 detected by mitoplasting and protease protection assay, figure has been modified from Wang et al., 201411. The total (T), pellet (P), and supernatant (S) fractions were subjected to western blotting analysis with indicated antibodies. TIMM13 (an inter-space protein), and HSP60 (a matrix protein). Please click here to view a larger version of this figure.
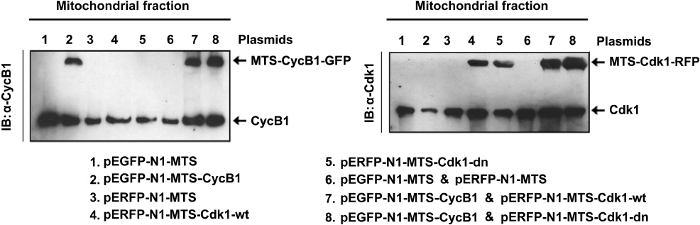
Figure 2. Expression of Mitochondrial Cdk1 Constructs. Western blotting of mitochondrial fractions isolated from cells transfected with mitochondria-targeted CyclinB1 and/or wild type or dominant negative mutant Cdk1 (plasmids are indicated on the bottom11. pEGFP-N1-MTS and the pERFP-N1-MTS vectors were empty vector controls for MTS-CyclinB1 and MTS-Cdk1 respectively). Please click here to view a larger version of this figure.
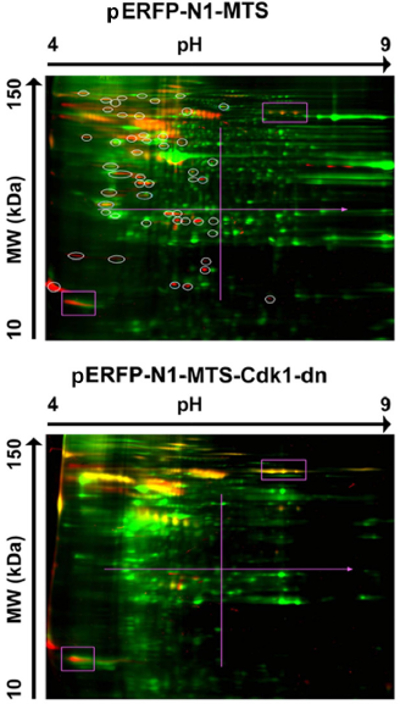
Figure 3. Potential Mitochondrial Substrates of Cdk1. Mitochondrial proteins extracted from G2/M-peaked cells transfected with mitochondria-targeted empty vector (pERFP-N1-MTS, upper panel) or mutant Cdk1 (pERFP-N1-MTS-Cdk1-dn, lower panel) were labeled with Cy5 (green), separated by 2-D gel and phosphorylated proteins were stained with phosphoprotein dye (red). This figure has been modified from Wang et al. 201411. Please click here to view a larger version of this figure.
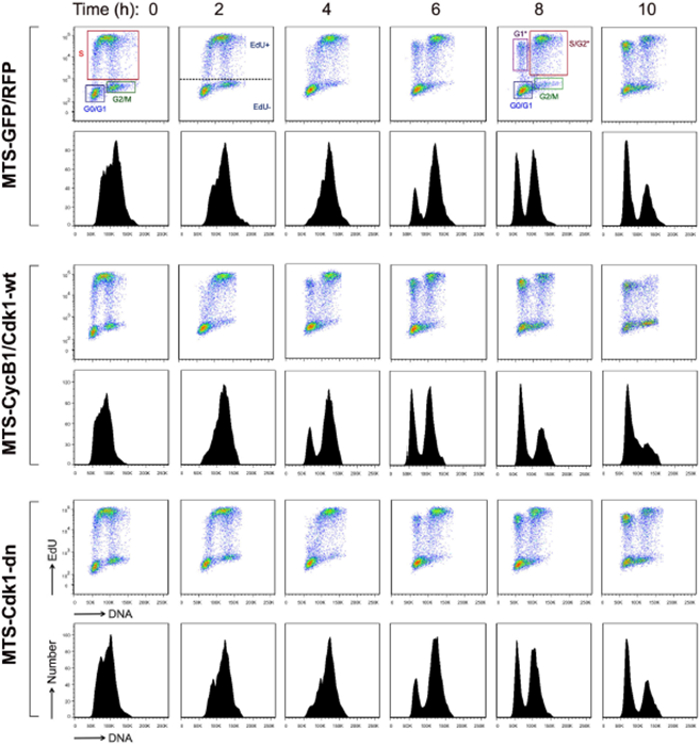
Figure 4. Mitochondrial Cdk1 Enhances G2/M Transition and Overall Cycle Progression.
Cell cycle analysis with EdU pulse-chase labeling. Scatter plot histograms of EdU-labeled cells were drawn for DNA content (X-axis) and EdU (Y-axis). The lower figures in each panel show the mean fluorescence intensity of the EdU labeled nuclei. The time points were indicated in h after the EdU pulse11. For all time points, gates displaying the following populations were drawn: G0/G1, S, and G2/M. For 6, 8, and 10-hr time points, EdU- labeled G1*, S/G2*, and G2/M* populations are shown. This figure has been modified from Wang et al., 201411. Please click here to view a larger version of this figure.
| Sucrose Concentrations Used | |||||
| No trypsin | 25 mM | 50 mM | 100 mM | 150 mM | 200 mM |
| + trypsin | 25 mM | 50 mM | 100 mM | 150 mM | 200 mM |
Table 1. Hypotonic Sucrose Buffers Used for Step 4.2
| Step 1 | 30 V | 12 hr | Step and Hold |
| Step 2 | 300 V | 0.5 hr | Step and Hold |
| Step 3 | 1,000 V | 0.5 hr | Gradient |
| Step 4 | 5,000 V | 1.33 hr | Gradient |
| Step 5 | 5,000 V | 20,000 V hr | Step and Hold |
Table 2. Isoelectric Protocol Used for Step 6.4.5