Male and female C57Bl/6J mice were euthanized on postnatal days 0-4 and the truncal skin was subjected to mechanical dissociation as described above. After chopping, the skin formed a viscous slurry lacking any sign of structural tissue. Centrifugation of this slurry resulted in the formation of a large cell pellet at the bottom of the conical tube and a layer of adipose floating on top of the supernatant. This adipose layer was discarded with the supernatant while the remaining cell pellet was resuspended and transferred to an uncoated well of a 6-well dish. The high density of cells caused the media to appear cloudy. However, after 40 min of incubation, larger cell and tissue conglomerates were observed in the media and adherent fibroblasts could be seen with an inverted light microscope (Figure 1).
The non-adherent cells from the fibroblast culture were transferred to one well of a collagen-coated 6-well dish in order to isolate melanocytes. Addition of G418 killed any remaining fibroblasts in the homogenate, leaving numerous dead cells floating in the media for the next 5 days (Figure 2A-C). After 4-5 days of growth, the plated melanocytes began to take on a stereotypical dendritic phenotype with melanocytic granules (Figure 2C). This phenotype persisted after passaging of the culture (Figure 2D). While clusters of contaminating keratinocytes were initially observed in the melanocyte cultures (Figure 2A), these populations were lost upon subsequent passaging.
Flow cytometric analyses were used to confirm the purity of the resulting primary melanocyte and fibroblast cultures. C5N mouse keratinocytes17, day 10 primary melanocyte cultures and day 6 primary fibroblast cultures were stained with: anti-gp100 (differentiated melanocytes), anti-Cytokeratin 14 (K14; keratinocytes) and anti-Fibroblast-specific protein 1 (FSP1; fibroblasts) (Figure 3). In these cells, gp100 staining was specific to the primary melanocytes, while FSP1 positivity was only observed in fibroblasts (Figure 3A,B). Expression of K14 was limited to C5N keratinocytes (Figure 3C). Additional analyses were performed to determine the purity of our melanocyte cultures 1, 2, 3, 4 and 10 days after isolation (Figure 4). Combined fibroblast and keratinocyte contamination never exceeded 25% of the total cells in culture (Figure 4B,C). Nevertheless, dramatic increases in gp100 were not observed until the fourth day in culture (Figure 4A). We attribute this observation to the reduced expression of gp100 in melanocyte precursors (i.e., melanoblasts), many of which appear fully differentiate by day 10 in culture (Figure 4A). In summary, these data demonstrate that our rapid isolation protocol simultaneously produces cultures enriched for melanocytes and fibroblasts.
| Step # | Reagent | 1 pup | 5 pups |
| 1.1 | Collagen Solution | 1.5 mL | 4.5 mL |
| 1.2 | Plate size | 6-well | 10 cm |
| 1.4 | Antibiotic / Antimycotic Solution | 3 mL | 15 mL |
| 1.5 | Skin Digestion Buffer | 3 mL | 15 mL |
| 1.6 | Melanocyte Media | 6 mL | 30 mL |
| 1.7 | Fibroblast Media | 4 mL | 20 mL |
Table 1: Reagent preparation guide for different cohort sizes.

Figure 1: Representative images of primary murine fibroblast cultures. Shown are 20x images of fibroblasts immediately following isolation (A), as well as 24 (B) and 48 (C) h post-isolation. Please click here to view a larger version of this figure.

Figure 2: Representative images of primary melanocyte cultures. Shown are 20x images of melanocyte cultures 1 (A), 2 (B) and 4 days (C) post-isolation. Contaminating keratinocytes are indicated by the arrow in 'A'. (D) Representative image of primary melanocyte cultures after passaging. Please click here to view a larger version of this figure.
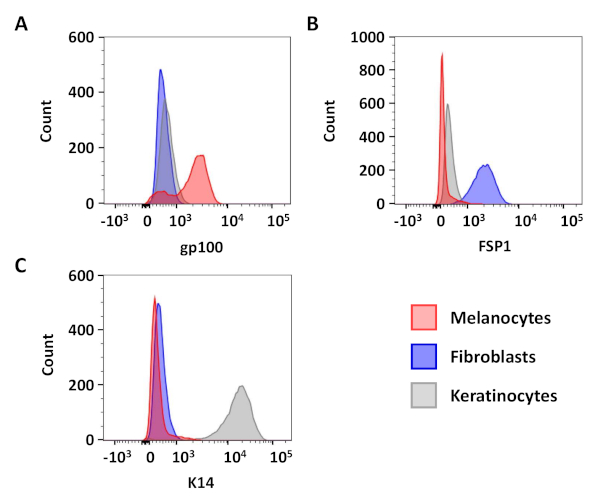
Figure 3: Flow cytometric assessments of culture purity. Representative histograms showing gp100 (A; differentiated melanocytes), FSP1 (B; fibroblasts) and K14 (C; keratinocytes) positivity in primary melanocytes (day 10), primary fibroblasts (day 6) and C5N keratinocytes. After gating on live cells, 10,000 events were analyzed from each population. Single color controls were used for compensation and the resulting data was visualized with FlowJo software. Please click here to view a larger version of this figure.
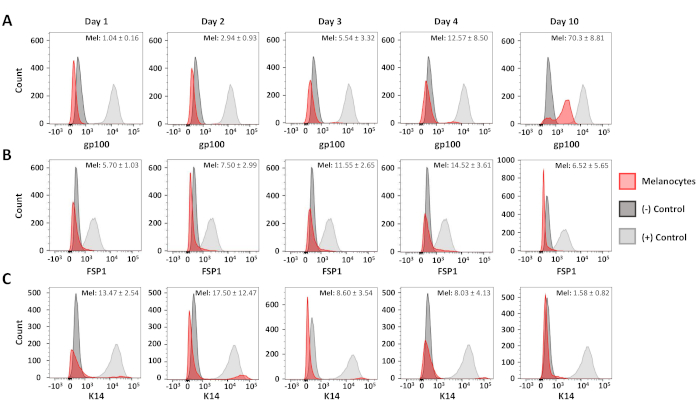
Figure 4: Time course analysis of melanocyte purity. Representative histograms showing melanocyte purity 1-10 days after initial isolation. Samples were stained, analyzed and graphed as described in Figure 3. Positive and negative control populations were as follows: gp100–human melanocytes (+), primary murine fibroblasts (-); FSP1–primary murine fibroblasts (+), human melanocytes (-); K14–C5N cells (+), human melanocytes (-). The average positivity and standard deviation for at least three distinct melanocyte cultures is shown in the upper right-hand corner of each graph. Please click here to view a larger version of this figure.