Adipogenesis is regulated by a network of transcription factors that are responsible for both the expression of key proteins that induce brown adipocyte formation and functioning22, including classical adipogenic regulators such as PPARγ and C/EBPα23,24,25, as well as markers of mature adipocytes26,27. Through testing the different concentrations of rosiglitazone that allow the acquisition of the thermogenic brown adipose phenotype, the method described herein allows the differentiation of preadipocytes present in the SVF of iBAT of newborn P0.5 mice to mature brown adipocytes. The addition of 5 µM rosiglitazone in the induction and maintenance medium markedly increased the protein levels of the adipogenic regulators and UCP1 at day 7 of differentiation compared with a lower concentration of this PPARγ agonist (1 nM) (Figure 3). Undifferentiated preadipocytes (day 0) present very low or undetectable levels of PPARγ, C/EBPα, PLIN1, and CD36. In contrast, these markers were significantly higher on day 7 of differentiation (Figure 4A–F). Consistent with the previously published results15, brown preadipocytes accumulated multiple lipid droplets across the differentiation protocol (Figure 5).
The mitochondrial mass increases during brown adipogenesis and is strongly associated with lipid droplet accumulation28. The present protocol markedly increased the mitochondrial mass marker TOM20 (Figure 6) and the OXPHOS complex (Figure 7) protein levels at day 7 of differentiation. UCP1 is highly expressed in mature brown adipocytes and is a bona fide marker of this cell type29. As shown in Figure 6, UCP1 was not detectable in undifferentiated preadipocytes (day 0) but was markedly increased at day 7 of differentiation.
Brown adipogenic differentiation is associated with changes in mitochondrial morphology and their physical association with other organelles15,30. As shown in Figure 8, the mitochondria evolved from an elongated tubular shape at day 0 toward a "rounded" or "bean-like" shape at day 7 (Figure 8A,B). The mitochondrial inner structure was also modified by adipogenesis, resulting in higher density of the parallel-packed cristae. Importantly, the mitochondria were intimately associated with lipid droplets in the differentiated brown adipocytes, so no discernible distance between the outer membrane of the mitochondria and the surfaces of the lipid droplets could be detected, even with high-resolution transmission electron microscopy (Figure 8C,D).
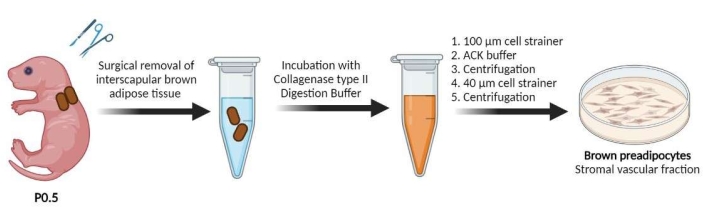
Figure 1: Generation of primary cultured preadipocytes from the SVF of the iBAT. Preadipocyte-containing SVFs were obtained from the interscapular adipose tissue of newborn (P0.5) mice. The interscapular brown adipose tissue was digested with collagenase type II digestion buffer. The blood cells were lysed with ACK buffer. The preadipocytes contained in the SVF were seeded in 24-well plates. Please click here to view a larger version of this figure.
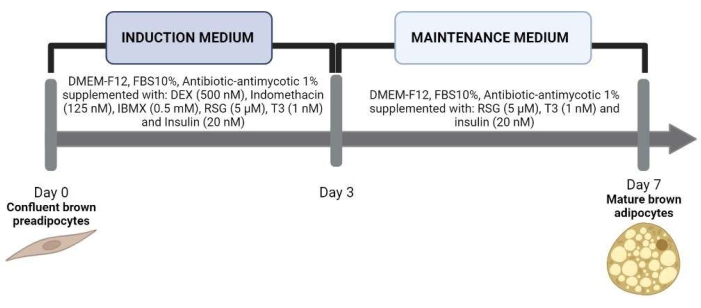
Figure 2: Induction of adipogenesis in primary cultured preadipocytes. Once the SVF cells reached confluency, adipogenesis was induced with the induction medium. On the third day of differentiation, this medium was replaced by a maintenance medium until day 7. Abbreviations: DMEM-F12 = Dulbecco's Modified Eagle Medium/Nutrient Mixture F-12; FBS = fetal bovine serum; DEX = dexamethasone; IBMX = isobutylmethylxanthine; RSG = rosiglitazone; T3 = triiodothyronine. Please click here to view a larger version of this figure.
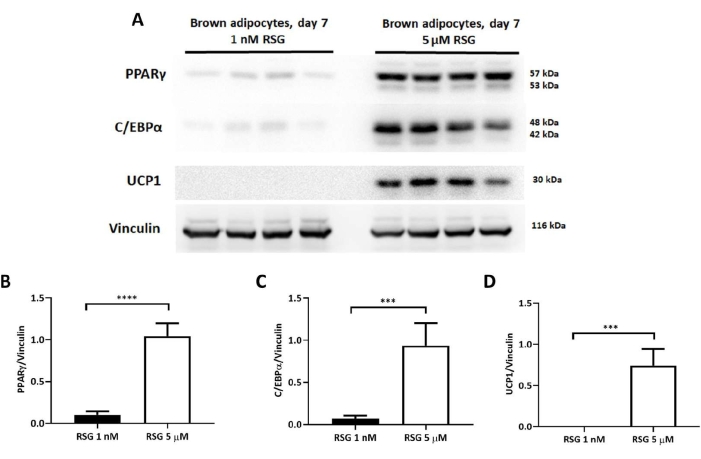
Figure 3: Protein levels of PPARγ, C/EBPα, and UCP1 in differentiated brown adipocytes differentiated with 1 nM or 5 µM rosiglitazone. (A) Representative immunoblot images of adipogenic regulators and UCP1 on day 7 of differentiation. Immunoblot quantification of (B) PPARγ, (C) C/EBPα, and (D) UCP1 protein levels; normalized to vinculin levels (n = 4 per experimental condition). Results expressed as mean ± SD. ***p < 0.001, and ****p < 0.0001 between adipocytes treated with 1 nM versus 5 µM rosiglitazone. The p-values were calculated using a Student's t-test. Please click here to view a larger version of this figure.
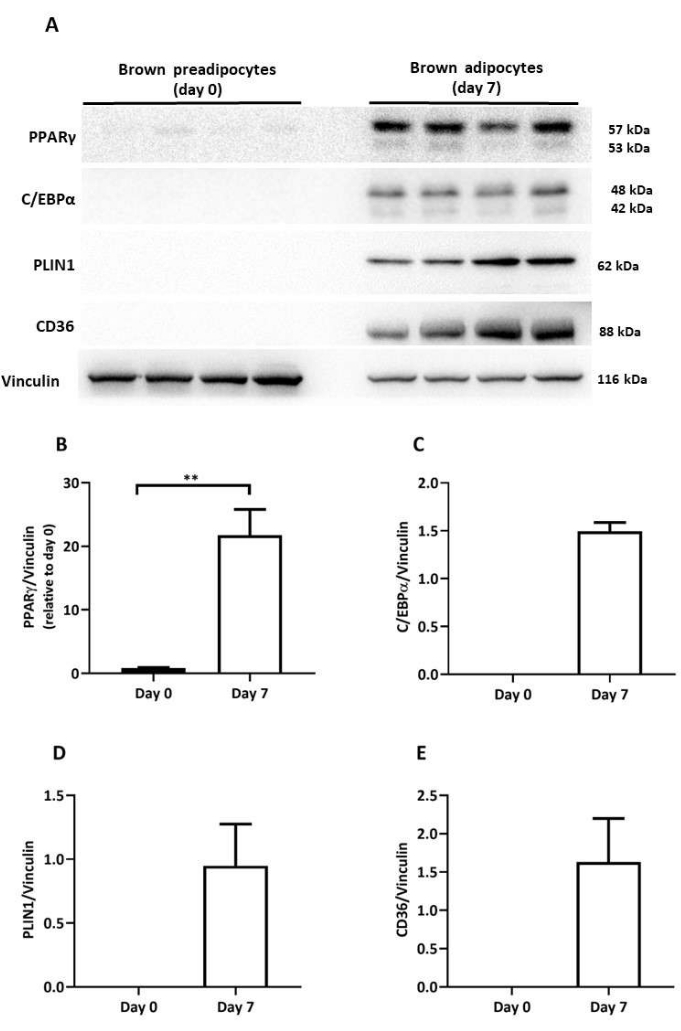
Figure 4: Protein levels of brown adipogenic regulators and mature adipocyte markers in differentiated brown adipocytes. (A) Representative immunoblot images of PPARγ, C/EBPα, PLIN1, and CD36of differentiated brown adipocytes on day 0 and day 7 of differentiation. Immunoblot quantification of (B) PPARγ, (C) C/EBPα, (D) PLIN1, and (E) CD36 protein levels; normalized to vinculin levels (n = 4 per differentiation day). Results expressed as mean ± SD. **p < 0.01 between undifferentiated (day 0) and differentiated brown adipocytes (day 7). The p-values were calculated using a Student's t-test. Please click here to view a larger version of this figure.
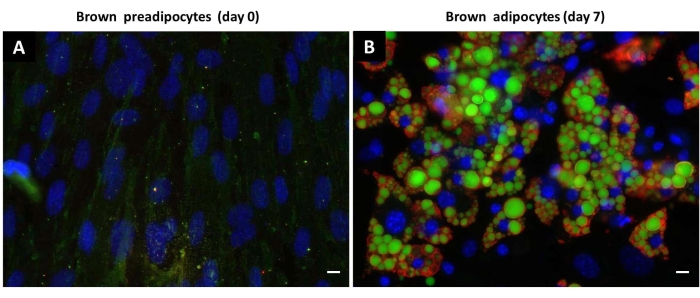
Figure 5: Accumulation of lipid droplets by differentiated brown adipocytes. Representative immunofluorescence images showing the staining of neutral lipids with BODIPY (green) and PLIN1 (red) and the staining of nuclei with Hoechst 33342 (blue) in brown (pre)adipocytes on day 0 and day 7 of differentiation. (A) Brown preadipocytes, day 0 of differentiation. (B) Brown adipocytes, day 7 of differentiation. Scale bar: 20 µm. Please click here to view a larger version of this figure.
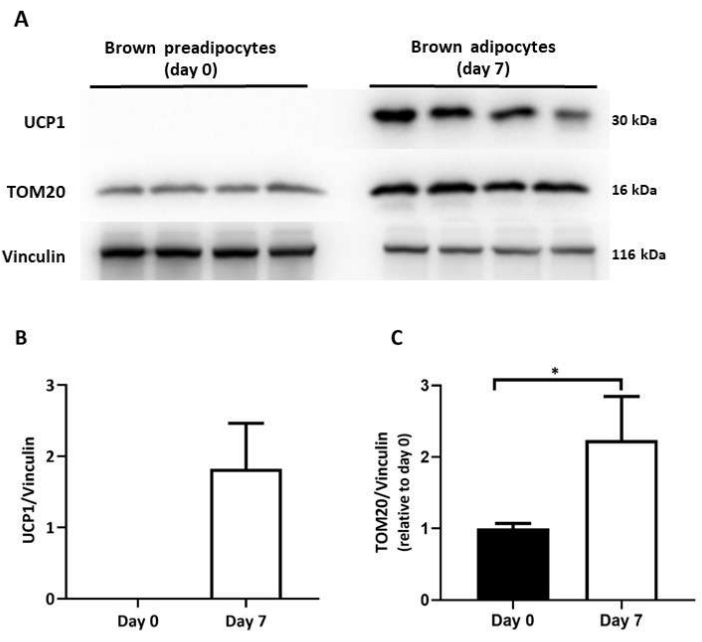
Figure 6: Protein levels of mitochondrial mass and thermogenic markers in differentiated brown adipocytes. (A) Representative immunoblot images of TOM20 and UCP1 on day 0 and day 7 of differentiation. Immunoblot quantification of (B) UCP1 and (C) TOM20 protein levels; normalized to vinculin levels (n = 4 per differentiaion day). Results expressed as mean ± SD. *p < 0.05 between undifferentiated (day 0) and differentiated brown adipocytes (day 7). The p-values were calculated using a Student's t-test. Please click here to view a larger version of this figure.
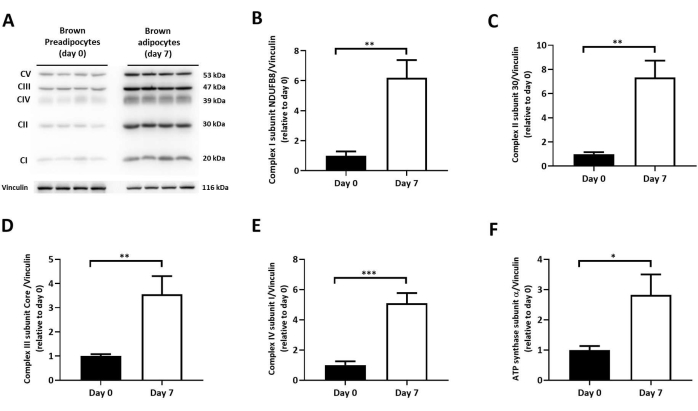
Figure 7: Protein levels of OXPHOS subunits in differentiated brown adipocytes. (A) Representative OXPHOS immunoblotting on day 0 and day 7 of differentiation. Immunoblot quantification of (B) complex I, subunit NDUFB8, (C) complex II, subunit 30, (D) complex III, subunit core, (E) complex IV, subunit I, (F) ATP synthase, subunit α; normalized to vinculin levels (n = 4 per differentiation day). Results expressed as mean ± SD. *p < 0.05, **p < 0.01, and ***p < 0.001 between undifferentiated (day 0) and differentiated brown adipocytes (day 7). The p-values were calculated using a Student's t-test. Please click here to view a larger version of this figure.
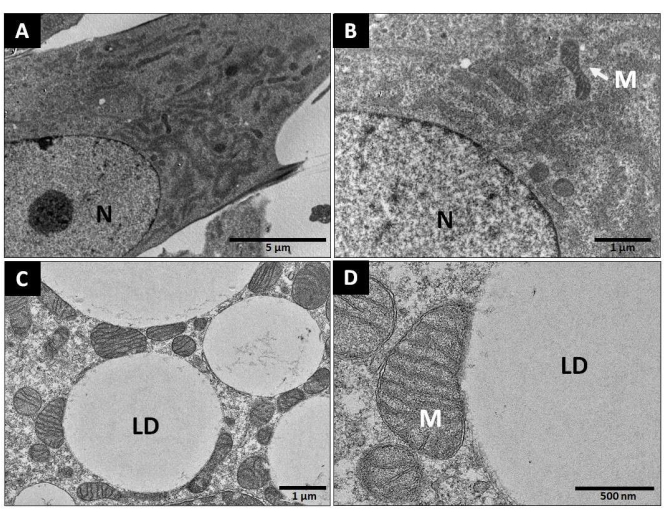
Figure 8: Mitochondrial morphology in differentiated brown adipocytes. Representative images of transmission electron microscopy on day 0 and day 7 of differentiation. (A,B) Brown preadipocytes on day 0 of differentiation; magnification: 4,200x and 16,500x, respectively. (C,D) Brown adipocytes on day 7 of differentiation; magnification: 8,500x and 28,000x, respectively. Abbreviations: LD = lipid droplet; M = mitochondria. Scale bar: (A) = 5 µm; (B,C) = 1 µm; (D) = 500 nm. Please click here to view a larger version of this figure.
Table 1: Buffer and medium preparation. Collagenase type II digestion buffer: Prepared in sterile deionized water, filtered, and aliquoted in a biosafety cabinet. For long-term storage, it is recommended to maintain it at −20 °C and thaw right before use. Culture medium: Adjusted to pH 7.2. For storage, it is recommended to maintain it at 4 °C. Induction and maintenance medium: Prepared in DMEM-F12, 10% FBS, 1% antibiotic-antimycotic, pH 7.2. Both these media must be freshly prepared just before use. Please click here to download this Table.
Supplementary Table 1: Comparison of the differentiation media used in various reported studies. Please click here to download this Table.
