

Summary
Dopamine vervanging farmacotherapie met L-dopa is de meest gebruikte symptomatische behandeling van Parkinson, maar gaat gepaard met bijwerkingen zoals onwillekeurige abnormale bewegingen, genoemd dyskinesie
Abstract
MALDI imaging massaspectrometrie (IMS) is een krachtige aanpak die de ruimtelijke analyse van moleculen in biologisch weefsel monsters 2 (fig. 1) vergemakkelijkt. Een 12 pm dunne weefselsectie is bedekt met een MALDI-matrix, die desorptie en ionisatie van intacte peptiden en eiwitten die gedetecteerd kan worden met een massa analysator, typisch met een MALDI TOF / TOF massaspectrometer vergemakkelijkt. Over het algemeen honderden pieken kan worden beoordeeld in een enkele rat hersenweefsel sectie. In tegenstelling tot de veelgebruikte beeldvormende technieken, is deze aanpak geen voorafgaande kennis van de moleculen van belang en maakt het mogelijk voor gebruik zonder toezicht en uitgebreide analyse van veelvoudige moleculaire species met behoud van een hoog moleculair specificiteit en sensitiviteit 2. Hier beschrijven we een MALDI IMS-gebaseerde aanpak voor het ophelderen van regio-specifieke verdeling profielen van neuropeptiden in de hersenen van de rat van de ziekte van een diermodel van Parkinson (PD).
PD is een veel voorkomende neurodegeneratieve ziekte met een prevalentie van 1% voor mensen ouder dan 65 en ouder 3,4. De meest voorkomende symptomatische behandeling op dopamine vervanging met L-DOPA 5. Maar dit gaat gepaard met ernstige bijwerkingen waaronder onvrijwillige abnormale bewegingen, genaamd L-DOPA-geïnduceerde dyskinesieën (LID) 1,3,6. Een van de belangrijkste moleculaire verandering in deksel een opregulatie van het opioïde precursor prodynorphin mRNA 7. De prodynorfine peptiden moduleren neurotransmissie in hersengebieden die voornamelijk betrokken zijn bij de beheersing van de bewegingen 7,8. Echter, om de exacte opioïde peptiden die afkomstig zijn van verwerking van de neuropeptide voorloper op heden niet bepaald. Daarom hebben we gebruikt MALDI IMS in een diermodel van de ziekte van Parkinson experimentele en L-dopa geïnduceerde dyskinesie.
MALDI imaging massaspectrometrie bleek bijzonder gunstig met betrekking tot neuropeptide karakteriseringTIE, omdat veel gebruikte antilichamen gebaseerde aanpak richt zich op bekende peptide sequenties en eerder waargenomen post-translationele modificaties. In tegenstelling MALDI IMS kan ontrafelen nieuwe peptide verwerking van producten en dus onthullen nieuwe moleculaire mechanismen van neuropeptide modulatie van neuronale transmissie. Hoewel het absolute bedrag van neuropeptiden niet kan worden bepaald door middel van MALDI IMS, de relatieve overvloed van peptide-ionen kan worden afgebakend van de massa spectra, het geven van inzicht over het wijzigen van het niveau bij gezondheid en ziekte. In de voorbeelden hier gepresenteerd, werden de piek intensiteiten van prodynorfine B, alfa-neoendorphin en substance P gevonden aanzienlijk worden verhoogd in de dorsolaterale, maar niet de dorsomediale, striatum van de dieren met ernstige dyskinesie met gezicht, romp en orolingual spieren (afb. 5). Bovendien, MALDI IMS bleek een correlatie tussen dyskinesie ernst en de mate van des-tyrosine alfa-neoendorphin, wat neerkomt op een voorheen onbekend mechanisme van functionele inactitie van dynorphins in het striatum het verwijderen van N-terminale tyrosine vermindert de prodynorfine de opioïde-receptor bindende capaciteit 9. Dit is de eerste studie over neuropeptide een karakterisering van het deksel met MALDI IMS en de resultaten benadrukken de mogelijkheden van de techniek voor toepassing in alle gebieden van het biomedisch onderzoek.
Protocol
Het protocol wordt aangepast ten behoeve van statistische analyse van MALDI IMS gegevens uit meerdere hersenen van de rat secties, meestal 20 tot 30 delen, en bestaat uit vijf verschillende stappen bestaande uit het prepareren van weefsels, matrix toepassing, MALDI-TOF MS-analyse, data-analyse, en neuropeptide identificatie. De procedures zijn beschreven en beschreven in meer gedetailleerde hieronder:
1. Tissue voorbereiding
Deze procedure omvat de verzameling van de respectieve weefsel alsmede weefsel snijden voor IMS analyse. Een bijzonder doel eiwitten en peptiden analyse te voorkomen proteolytische afbraak. Daarom is het essentieel om snel en zorgvuldig te werken tijdens weefsel dissectie.
- Offer ratten (250-300 g typisch) werden onthoofd verwijderen rattenhersenen binnen maximaal postmortale van <30 en bevriezen poedervormig droog ijs alvorens tot -80 ° C diepvriezer. Sneller vriezen met vloeibare stikstofverhogen het risico van microtears in het hersenweefsel, die negatief zal beïnvloeden matrix kristallisatie en daardoor niet MS kwaliteit (fig. 2D). Hele hersenen kunnen worden opgeslagen voor een aantal jaren voordat het snijden zonder verlies van MS kwaliteit van het signaal.
- Snijd bevroren weefsel op een cryostaat microtoom tot 12 micrometer plakjes en dooi-mount weefselcoupes op een geleidende MALDI glazen dia's (indium tin oxide coating dia's, Bruker Daltonics) of MALDI doel (afb. 2A-C).
- Droog secties 15 min onder vacuüm en opslaan dia bij -80 ° C tot verder gebruik. Weefselsecties worden geanalyseerd in de kortst mogelijke tijd na snijden, zelfs indien bewaard bij -80 ° C. We vinden dat MS kwaliteit van het signaal zal merkbaar worden verminderd na een jaar in opslag. Om oxidatie van eiwitten en peptiden verminderen, kan de lucht in opslagvat worden vervangen door een inert gas (bijv. argon of stikstof).
2. Matrix toepassing
De matrix applicatiestap heeft een aanzienlijke invloed op de kwaliteit spectrum en vereist optimalisatie van verschillende parameters afhankelijk van het soort weefsel en de analyt van belang. Deze factoren omvatten chemische parameters zoals de aard van de matrix, matrix concentratie, pH, weefsel wassen en organische modificatoren en instrumentele instellingen met borg volume laterale resolutie en het aantal van afzettingen 10 (Fig. 2D). Voor grote schaal experimenten is het van groot belang variantie verminderen, bijvoorbeeld door toepassing van de matrix voor alle lagen binnen een dag door dezelfde. Hoewel er vele strategieën matrix oplossing te brengen zoals door sublimatie of spray, het automatisch aanbrengen van reeksen van kleine druppeltjes matrix, ongeveer 100-150 picoliter groot is met succes gebruikt voor de analyse van kleine eiwitten en peptiden in verschillende weefsels , waaronder hersensecties 9, 10,11, 12, 13.
- Ontdooi de secties in een exsiccator gedurende 1 uuronze.
- Zorg ervoor dat het experiment wordt verblind door een andere persoon dan de exploitant. Re-label alle monsters.
- Was secties 1x in 70% ethanol (EtOH, bij kamertemperatuur, RT) 10 sec en tweemaal in 95% EtOH (RT) 10 sec. Voor grote experimenten, voer dan de wassen voor alle glas schuift samen met een cuvet om variatie te minimaliseren.
- Droog de secties in een exsiccator gedurende 10 minuten.
- Evalueer weefselcoupes onder een microscoop en controleer weefsels vervorming, microtears en kleine scheuren die zal schaden MALDI MS kwaliteit (fig. 2D).
- Bereid matrix oplossing bestaande uit 50 mg / ml DHB in 50% methanol, 10% 150 ammoniumacetaat (Amac) en 0,3% trifluorazijnzuur (TFA) in water.
- Matrix toepassing door discrete druppels afzetting in een rechthoekig patroon met een chemisch inkjetprinter (CHIP, Shimadzu). De eerste stap is om de experimentele parameters van matrix toepassing te optimaliseren voor het neuropeptide analyse including van het aantal druppels per pas, aantal passages. Dit experiment wordt uitgevoerd door het toepassen van meerdere matrix arrays van verschillende toepassingen met parameters op dezelfde weefselsectie, terwijl het ervoor zorgen dat elke array soortgelijke gebieden van de hersenen, zoals corpus callosum, cortex en het striatum bekleding. Hetzelfde experiment wordt uitgevoerd telkens parameters worden gewijzigd, met inbegrip van verschillende hersenstructuren verschillende matrices voor specifieke analyten, en indien verschillende matrix oplosmiddelen nodig zijn voor winning van specifieke analyten.
- Scan het glaasje houder met de weefselsectie en lijn de houder. Definieer uw array voor matrix toepassing op de weefselsectie en geef de ruimtelijke resolutie dat wil zeggen plek om afstand te spotten. Breng matrix met behulp van de geoptimaliseerde protocol over de chemische inkjet printer. Voor dit experiment hebben we een geoptimaliseerd protocol voor peptide beeldvorming met de volgende drukkerij parameters gebruikt: 10 druppels (100 PL / drop), 10 toepassing passeert en een plek om te zien distance van 300 urn.
- Scan laatste matrix gespot secties en de afbeelding opslaan voor registratie voorafgaand aan de MALDI data-acquisitie (stap 3.4).
- Bewaar de profielen tot gebruik in een exsiccator onder vacuüm.
3. MALDI MS data-acquisitie en-verwerking
MS analyse van neuropeptiden wordt uitgevoerd op een MALDI time of flight instrument (Ultraflex II, Bruker Daltonics, Duitsland) die actief zijn in reflector-modus, met behulp van software ondersteunde data-acquisitie van elke matrix plek 14. Daarom is nauwkeurige ruimtelijke onderwijs is noodzakelijk. Het is essentieel dat de MALDI optimalisatie verkrijging vooral het doel registratie experimenten uitgevoerd door dezelfde die bij voorkeur worden blind de experimentele groepen. In een grootschalig experiment met meerdere glazen dia's, kan de MALDI experimenten worden uitgevoerd door een operator, terwijl een andere persoon is het gebruik van de chemische inkjet printer.
- Laad glazen dia's in de massaspectrometer.
- Controleer de ijking van de MALDI overnamemethode met een laag molecuulgewicht standaardijking mix (Bruker Daltonics).
- Optimaliseer acquisitie parameters.
- Om MS signaal te optimaliseren en voorkomen wegnemen matrix van naburige matrix afzettingen, moet de grootte van de laser en de optimale aandacht weefsel te bepalen.
- De laser-energie is ingesteld op de maximale MS kwaliteit te waarborgen van zoveel matrix deposito's mogelijk zonder het heffen van de baseline, waardoor de piek resolutie of het verzadigen van de detector.
- Beoordeel het maximum aantal opnamen per matrix plek tot er alleen nog geluid wordt gedetecteerd, vaak 1000-2000 schoten. Schat het aantal opnamen die moeten worden verzameld en het aantal foto's die vóór de laser positie binnen een plek zou moeten veranderen. Om elke matrix plaats gelijkmatig proeven, verzamelen we 600 opnamen in 25 shot stappen een totaal aantal van 24 stappen met een random bewegingspatroon van elke matrix depositie.
- Registreer de scan van alle gevlekte onderdelen van de motor coördinaten van de MALDI stadium met behulp van de FlexImaging (v.2.0) 10 en het uitvoeren van data-acquisitie in batch modus door AutoXexuteBatchRunner.exe software.
- Verwerk elke afzonderlijke spectra door middel van baseline aftrekken (convex omhulsel V3), smoothing en externe kalibratie (optioneel), gevolgd door export als een ASCII-bestand (*. Dat, *. Txt of *. Csv-formaat). 15
4. Evaluatie van de gegevens
Definitieve data evaluatie bevat gegevens na verwerking en data reductie door richten zich uitsluitend op piek informatie, gevolgd door een statistische analyse.
- In een eerste stap werden de MALDI IMS secties geëvalueerd overnormalization effecten. Dit kan gemakkelijk worden bereikt door het gebruik van data visualisatie tools zoals FlexImaging (Bruker Daltonics) of BioMap (Novartis). In een eerste fase de totale ionen beelden zijn evaluated voordat de totale ion huidige (TIC) normalisatie, gevolgd door handmatige inspectie van enkele ionen distributie afbeeldingen van diverse prominente peptide pieken. Kijk voor karakteristieke piek intensiteit distributies en als ze gerelateerd zijn aan weefsel kenmerken (schade), het spotten van de kwaliteit of de normalisering effecten (afb. 3).
- Af te bakenen regio's van belang (bijvoorbeeld het striatum) op basis van histologische kenmerken en exporteren van de bijbehorende spectra in ASCII-formaat. Bij voorkeur kunnen de normalisatie van spectra van de totale ionenstroom (TIC) omvat in dit stadium.
- ASCII-bestanden in een data-handling software zoals Origin (v.8.1, Originlab), MATLAB (MathWorks, Natick, MA, Verenigde Staten) of R 16. Peak detectie kan worden uitgevoerd met behulp van peak het vinden van tools in de software, bijvoorbeeld "piek-analyse" in Origin of "mspeaks" in Matlab. Exporteer de peaklists van alle spectra als een enkele tab gescheiden tekstbestand.
- Met het oog op bin grenzen te bepalen voor gedetecteerd peptidepieken, wordt binning analyse uitgevoerd met de juiste software tools (bijvoorbeeld pbin 17) of in-house geschreven scripts voor MATLAB of R. Hier is de tekst bestand met alle piek geplukt data wordt geladen in de software en de parameters voor piek-grens bepaling worden gespecificeerd zoals hoe vaak een piek moet in spectra om relevant zijn voor de proef. Bijvoorbeeld, de experiment bevat twee groepen dieren, 5 dieren in elke groep en 100 spectra worden verzameld per dier en omgeving van belang. Neem aan dat een piek is in potentie interessant als het aanwezig is in ten minste de meerderheid van de dieren in een groep (3/5) en in ten minste de helft van de spectra van deze dieren (3x50 = 150 spectra), zal dit een totaal percentage van 15% voor de 150 positieve spectra van de in totaal 1000 (2x5x100) spectra. Met behulp van de pbin tool, deze stap levert een binrange bestand met alle bin breedtes bepaald op basis van de verkregen gegevens. Om te voorkomen dat bin de grenzen te controlerengeschikt is eenvoudig samen te visualiseren van de bakken in oorsprong het oorspronkelijke spectra sporen.
- Peak-area integratie kan verminderen variantie die van belang is voor statistische analyse. We maken gebruik van een eigen geschreven script voor R om de oppervlakte te berekenen onder de curve tussen piek-grenzen bepaald in stap 4. Geïntegreerde piekoppervlakken worden geïmporteerd in MS Excel (v.2007) en statistische analyse door middel van niet-parametrische ongepaarde testen met behulp van de SAM-programma wordt uitgevoerd 18.
5. Peptide identificatie
Volgorde verificatie van de waargenomen peptide identiteiten is van essentieel belang om biologische relevantie te sluiten. De meest nauwkeurige methode is onder andere echte top-down bepaling direct van weefsel met behulp van peptide fragmentatie door middel van tandem massaspectrometrie (MS / MS), hoewel de hoge peptide concentraties nodig zijn voor dit soort analyses 12,13. Voor lage overvloedige peptiden of meerdere peptiden met closem / z-waarden (± 0,5%), wordt op weefsel analyse aangetast en uit weefsel analyse met behulp van een peptidomic strategie wordt gebruikt, dat omvat de extractie, scheidings-en MS-gebaseerde identificatie van endogene neuropeptiden. Voor experiment hier gepresenteerde centraal was opioïde peptide detectie is een bijzondere uitdaging omdat deze peptiden zeer laag vergeleken met andere overvloedig neuropeptiden in de spectra. Bovendien zijn deze peptiden vrij polaire waardoor ze relatief hydrofiele en moeilijk te handhaven met gemeenschappelijke peptide extractie en scheidingstechnieken .. Daarom hebben wij gebruik gemaakt van een eerder gemeld protocol voor weefsel-extractie en opioïde peptide voorfractionering in combinatie met standaard LC-MS/MS gebaseerd peptide identificatie 9,19.
- Verzamel coronale secties van target structuren van belang (nucleus accumbens, NAC, nucleus caudatus putamen, CPU). Monteer bevroren hersenen van de rat in een cryostaat microtoom en verwijder omliggende hersenmateriaal (cortex, septum, corpus callosum) met een scalpel. Verzamel secties (30 micrometer; n = 50) van de ontleed NAC en CPU en dooi monteer de NAC-en CPU-delen van de onderwerpen op verschillende glazen dia's.
- Extract peptiden uit weefsel door toevoeging van 100 ul 5% ACN/0.1% TFA, incubeer gedurende twee minuten en verzamel in Eppendorf lage eiwitbinding buizen. Herhaal deze stap twee keer.
- Voer peptide voorfractionering met sterke kationenuitwisselingschromatografie met stapsgewijze (n = 4) elutie verhoogd ionensterkte 19. Droog naar beneden monsters onder vacuüm met behulp van een speedvac concentrator.
- Analyseer de peptide fracties door middel van nanoflow C18 reversed phase vloeistofchromatografie (1100, Agilent Technologies, Santa Clara, CA) gekoppeld aan electrospray tandem massaspectrometrie (LC-MS/MS). De MS experimenten werden uitgevoerd op een hybride lineaire iontrap / Fourier transform ion cyclotron resonantie (FTICR) instrument (LTQ FT 7T, Thermo Scientific, Waltham, MA,) Peptide fulscan spectra (m / z 150-2.000) Werden verworven met de FTICR analyser met hoge massa resolutie (100.000), gevolgd door verdere versnippering van de 5 meest intense peptide pieken in de iontrap door middel van botsing geïnduceerde dissociatie (CID) 9.
- Peptidensequentie identificatie wordt uitgevoerd door database-matching en kan worden aangevuld met de-novo sequentie-analyse. Voor de database, worden in de handel verkrijgbare zoekmachines (Mascot, XTandem of Protein goudzoeker) in dienst 20. De zoekopdrachten worden meestal uitgevoerd tegen de databases met sequenties van bekende of voorspelde neuropeptiden en sequenties van neuropeptide precursor eiwitten 21.
6. Representatieve resultaten
MALDI beeldvorming massaspectrometrie van striatale weefselcoupes zoals bereid volgens het protocol beschreven resulteerde in de van meer dan 1000 pieken overeenkomt met ongeveer 300 mono-isotopische moleculaire (gemiddelde spectra weergegevenin Fig. 1). Data visualisatie voor prominente moleculaire ion piek werd bereikt met behulp van de Flex Imaging software en toonde karakteristieke piek intensiteit distributies die goed in lijn zijn met anatomische kenmerken (fig. 3). Een ander kenmerk van MALDI IMS is de relatieve goede reproduceerbaarheid. In dit experiment de totale variatiecoëfficiënt de piekintensiteiten van alle gedetecteerde moleculaire was 30%, maar veel pieken weergegeven zeer geringe variatie hoge reproduceerbaarheid binnen behandelde groepen (Fig. 4). De relatieve piek van de lichtsterkte afhankelijk van vier verschillende regio's van belang, met inbegrip van de dorsolaterale en dorsomediale deel uit van zowel de gelaedeerde en intact striata werden onderworpen aan statistische analyses. Om te corrigeren voor verschillende vergelijkingen simultaan werd de statistische analyse door middel van parametrische getest volgens de SAM gereedschap 18. De meest prominente veranderingen werden gevonden in de dorsolaterale deel van de dopamine-gedenerveerde, Parkinson striatum. Hier si lijke veranderingen teweegbrengen veranderingen in tussen de verschillende behandelingsgroepen werden waargenomen voor twee prodynorfine peptiden; prodynorfine B en alfa-neoendorphin (figuur 5). In detail, een relatieve toename van zowel prodynorfine intensiteit van de piek met 50-60% werd waargenomen bij hoge dyskinetische dieren in vergelijking met lage dyskinetische dieren en laesie controles (p <0,05, F (2, 15) = 12,8 DynB, F = 5,7 aNeo; Fig. 5).

. Figuur 1 Gemiddelde MS traceert verkregen uit twee nauw met elkaar verbonden regio's van het striatum, caudate putamen (CPU) en de nucleus accumbens (NAC). De twee regio's weer te geven verschillende MS-profielen met een aantal moleculen op unieke uitgedrukt in een regio, of op verschillende piekintensiteit niveaus (insert, m / z 2028). De ruimtelijke verdeling patroon van elke piek kan worden gevisualiseerd met behulp van gespecialiseerde beeldbewerkingssoftware (onderste paneel).
2.jpg "/>
Figuur 2 (A) De hersenen gemonteerd op een cryostaat boorkop met een verankering media (OTC, pijl). Wordt gezorgd dat de OTC niet het gebied van hersenen doorsnede zijn omdat de OTC oorzaak ion onderdrukking van peptiden besmetten. (B, C) Slijpplaten (≈ 12 micrometer dikte) zijn ontdooien gemonteerd op MALDI compatibel glasplaatjes en gedroogd enkele seconden vorstschade vermijden gezien C. (D) Microtears kan moeilijk te detecteren met het blote oog , maar aantasting MALDI matrix kristallisatie en vernietigen MALDI MS signaal. Hetzelfde deel gekleurd met cresyl violet onthult microtears en scheuren (rechtsonder microfoto).

Figuur 3. De eerste stap in de evaluatie van gegevens is een aantal verschillende pieken te visualiseren over de massabereik geanalyseerd (AI). Hier werden striatale secties van 9 muizen afgebeeld met MALDI MS. Visualisatie van de gemiddelde totale iop de huidige zal onthullen gebieden met opvallend hoge of lage ionenintensiteiten (pijlen). Deze gebieden kunnen worden beïnvloed door over-of onder-normalisatie-effecten en verstoren data-analyse te doen aan de resultaten. Slechte anatomische definitie van piek-distributies laten zien secties met over het algemeen lage piek signaal-ruis, bijvoorbeeld delen 3 en 9, pieken F tot en met I.

Figuur 4. MS reproduceerbaarheid tussen de groepen, kan worden beoordeeld door berekening van de gemiddelde MS-trace en de standaardfout voor iedere m / z-waarde (inserts, m / z 722 en 1749). Goede reproduceerbaarheid zorgt geldige statistische analyse.
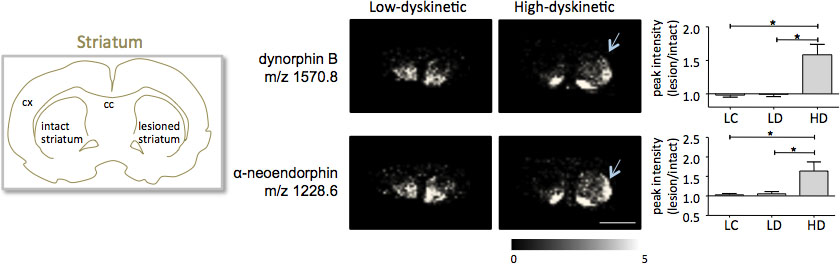
Figuur 5. Prodynorfine B en alfa-neoendorphin piek intensiteiten zijn sterk toegenomen in de 6-OHDA-Gelaedeerde, Parkinson, striatum van high-dyskinetische dieren (HD; pijlen) in vergelijking met de lage-dyskinetische (LD) en laesie controle groep (LC). Peptide piek intensiteiten uitgedrukt als gemiddelde fold-verandering van de intacte kant ± SEM (laesie / intacte zijde). * P <0,05; cx cortex; cc corpus callosum. Schaal bar 5 mm.
Discussion
Er zijn verschillende voordelen van de tewerkstelling van MALDI imaging massaspectrometrie in de studie van neuropeptiden. Een zuivere analyse van MS gegevens blijkt dat slechts bepaalde hersenkernen, of in de hier gepresenteerde resultaten waarin alleen de dorsolaterale deel van het striatum verbonden met een bepaalde pathofysiologische toestand. Door het behoud van de ruimtelijke informatie is het dan mogelijk om gebieden die van belang te herdefiniëren om statistische analyses uit te voeren met een hogere gevoeligheid en een lagere variabiliteit in vergelijking met de analyse van het hele brein secties of met behulp van traditionele peptidomics studies over peptide extracten. Bovendien is het belangrijk om te realiseren MALDI IMS gemakkelijk kan onbekende post-translationele modificaties detecteren, maar structurele analyse moet volgen om het exacte aminozuurposities die worden gewijzigd bepalen.
Voorkomende valkuilen in het visualiseren van de MALDI IMS gegevens op te nemen in kaart brengen van de maximale piek intensiteit naar een lineaire optische schaal van black (0%) de kleur (100%) voor elke sectie in de reeks experimenten (figuur 3), in plaats van in kaart brengen alle secties een gemeenschappelijke absolute schaal waarbij 100% van de maximale piek-intensiteit van alle secties (figuur 5) . Deze laatste methode maakt het mogelijk de vergelijking van de groep de gegevens en visualisatie van de verschillen tussen de behandelingsgroepen.
Een belangrijk obstakel in de MALDI IMS analyse is de toewijzing van peptiden aan soortelijke massa pieken. On-weefsel tandem-massaspectrometrie is het soms mogelijk, maar vaak blijkt heel moeilijk 13,14. We merken dat een meer traditionele benadering met een preparatieve fractionering op sterk kationenuitwisselingschromatografie, gevolgd door reversed phase LC-MS/MS kan worden met succes sequentie vele neuropeptiden en vooral opioïde peptiden. Het is nog steeds niet ongewoon om het verkrijgen van goede kwaliteit MS / MS-spectra die niet overeenkomen met een database gegevens het gebruik van gemeenschappelijke zoekmachines als mascotte. In deze gevallen de novo-sequencing hand de only optie. Het ultieme bewijs van de piek identiteit kan worden verkregen door MALDI IMS van weefselcoupes van de juiste knockout muis, maar dit is niet altijd beschikbaar of mogelijk is. Een alternatief is het valideren met een diametraal andere methode, bijvoorbeeld door Western immunoblotting of immunohistochemie. Dit kan bevatten vaak opwekken van antilichamen en veel werk valideren nieuwe antilichamen.
De algemene strategie die in dit protocol is geoptimaliseerd voor grootschalige neuropeptide MALDI IMS-experimenten, inclusief een aantal secties en experimentele omstandigheden. Het protocol is speciaal geoptimaliseerd voor de opioïde peptiden en zal grote impact hebben in toekomstige studies, zoals toegepast in diverse gebieden van onderzoek met inbegrip van de mechanismen die ten grondslag liggen aan pijn en de endogene reactie op drugs verslaving.
Disclosures
De auteurs hebben niets te onthullen.
Acknowledgments
Wij danken Hanna Warner om bij te dragen de gegevens voor figuur 3 en prof. Jonas Bergquist voor waardevolle input. De Zweedse Research Council (Grant 522-2006-6416 (MA), 521-2007-5407 (MA); De Åke Wiberg's Foundation (MA, JH), De Koninklijke Zweedse Academie van Wetenschappen (MA, JH), en de Zweedse Chemische Society (JH) worden bedankt voor financiële steun.
References
- Obeso, J. A., Olanow, C. W., Nutt, J. G. Levodopa motor complications in Parkinson's disease. Trends Neurosci. 23, S2-S7 (2000).
- Caprioli, R. M., Farmer, T. B., Gile, J. Molecular imaging of biological samples: localization of peptides and proteins using MALDI-TOF MS. MALDI-TOF MS. Anal. Chem. 69, 4751-4760 (1997).
- Obeso, J. A. The evolution and origin of motor complications in Parkinson's disease. Neurology. 55, S13-S20 (2000).
- O, W. H. Noncommunicable Diseases and Mental Health Cluster, Noncommunicable Disease Prevention and Health Promotion Department, Ageing and Life Course. Active Ageing: A Policy framework. , (2002).
- Schapira, A. H. Movement disorders: advances in cause and treatment. Lancet Neurology. , 6-7 (2010).
- Obeso, J. A., Rodriguez-Oroz, M. C., Rodriguez, M., DeLong, M. R., Olanow, C. W. Pathophysiology of levodopa-induced dyskinesias in Parkinson's disease: problems with the current model. Ann. Neurol. 47, S22-S32 (2000).
- Cenci, M. A., Lee, C. S., Bjorklund, A. L-DOPA-induced dyskinesia in the rat is associated with striatal overexpression of prodynorphin- and glutamic acid decarboxylase mRNA. Eur. J. Neurosci. 10, 2694-2706 (1998).
- Andersson, M., Hilbertson, A., Cenci, M. A. Striatal fosB expression is causally linked with l-DOPA-induced abnormal involuntary movements and the associated upregulation of striatal prodynorphin mRNA in a rat model of Parkinson's disease. Neurobiol Dis. 6, 461-474 (1999).
- Hanrieder, J. Alterations of striatal neuropeptides revealed by imaging mass spectrometry. Molecular & Cellular Proteomics. , (2011).
- Cornett, D. S., Reyzer, M. L., Chaurand, P., Caprioli, R. M. MALDI imaging mass spectrometry: molecular snapshots of biochemical systems. Nat. Methods. 4, 828-833 (2007).
- Ljungdahl, Imaging Mass Spectrometry Reveals Elevated Nigral Levels of Dynorphin Neuropeptides in L-DOPA-Induced Dyskinesia in Rat Model of Parkinson's Disease. PLoS ONE. 6, e25653 (2011).
- Groseclose, M. R., Andersson, M., Hardesty, W. M., Caprioli, R. M. Identification of proteins directly from tissue: in situ tryptic digestions coupled with imaging mass spectrometry. J. Mass. Spectrom. 42, 254-262 (2007).
- Andersson, M., Groseclose, M. R., Deutch, A. Y., Caprioli, R. M. Imaging mass spectrometry of proteins and peptides: 3D volume reconstruction. Nat. Methods. 5, 101-108 (2008).
- Deininger, S. -O. Imaging Mass Spectrometry. Setou, M. , Springer. Japan. 199-208 (2010).
- Norris, J. L. Processing MALDI Mass Spectra to Improve Mass Spectral Direct Tissue Analysis. Int. J. Mass. Spectrom. 260, 212-221 (2007).
- Ihaka, R., Gentleman, R. R. A Language for Data Analysis and Graphics. Journal of Computational and Graphical Statistics. 5, 299-314 (1996).
- Mass Spectrometry Binning Software GAB. , Vanderbilt Center for Quantitative Sciences. Nashville, TN. Available from: http://www.vicc.org/biostatistics/software.php (2012).
- Tusher, V. G., Tibshirani, R., Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. U.S.A. 98, 5116-5121 (2001).
- Bergstrom, L., Christensson, I., Folkesson, R., Stenstrom, B., Terenius, L. An ion exchange chromatography and radioimmunoassay procedure for measuring opioid peptides and substance P. Life. Sci. 33, 1613-1619 (1983).
- Falth, M. Neuropeptidomics strategies for specific and sensitive identification of endogenous peptides. Mol. Cell. Proteomics. 6, 1188-1197 (2007).
- Falth, M. SwePep, a database designed for endogenous peptides and mass spectrometry. Mol. Cell. Proteomics. 5, 998-1005 (2006).
