

ERRATUM NOTICE
Important: There has been an erratum issued for this article. Read more …
Summary
De neuron-gliale interacties bij neurodegeneratie worden niet goed begrepen als gevolg van ontoereikende hulpmiddelen en methoden. Hier beschrijven we geoptimaliseerde protocollen om geïnduceerde neuronen, oligodendrocytenvoorlopercellen en oligodendrocyten te verkrijgen uit menselijke pluripotente stamcellen en geven we voorbeelden van de waarden van deze methoden bij het begrijpen van celtypespecifieke bijdragen bij de ziekte van Alzheimer.
Abstract
Bij de ziekte van Alzheimer (AD) en andere neurodegeneratieve aandoeningen is oligodendrogliaal falen een veel voorkomend vroeg pathologisch kenmerk, maar hoe het bijdraagt aan de ontwikkeling en progressie van de ziekte, met name in de grijze stof van de hersenen, blijft grotendeels onbekend. De disfunctie van oligodendrocytenafstammingscellen wordt gekenmerkt door tekortkomingen in myelinisatie en verminderde zelfvernieuwing van oligodendrocytenvoorlopercellen (OPC's). Deze twee defecten worden ten minste gedeeltelijk veroorzaakt door de verstoring van interacties tussen neuronen en oligodendrocyten langs de opbouw van pathologie. OPC's geven aanleiding tot myeliniserende oligodendrocyten tijdens de ontwikkeling van het CZS. In de volwassen hersenschors zijn OPC's de belangrijkste proliferatieve cellen (bestaande uit ~ 5% van de totale hersencellen) en controleren nieuwe myelinevorming op een neurale activiteitsafhankelijke manier. Dergelijke neuron-naar-oligodendrocytencommunicatie wordt aanzienlijk onderbelicht, vooral in de context van neurodegeneratieve aandoeningen zoals AD, vanwege het gebrek aan geschikte hulpmiddelen. In de afgelopen jaren hebben onze groep en anderen aanzienlijke vooruitgang geboekt bij het verbeteren van de momenteel beschikbare protocollen om functionele neuronen en oligodendrocyten individueel te genereren uit menselijke pluripotente stamcellen. In dit manuscript beschrijven we onze geoptimaliseerde procedures, waaronder het opzetten van een co-kweeksysteem om de neuron-oligodendrocytenverbindingen te modelleren. Onze illustratieve resultaten suggereren een onverwachte bijdrage van OPC's / oligodendrocyten aan de amyloïdose van de hersenen en synapsintegriteit en benadrukken het nut van deze methodologie voor AD-onderzoek. Deze reductionistische benadering is een krachtig hulpmiddel om de specifieke hetero-cellulaire interacties te ontleden uit de inherente complexiteit in de hersenen. De protocollen die we hier beschrijven, zullen naar verwachting toekomstige studies naar oligodendrogliale defecten in de pathogenese van neurodegeneratie vergemakkelijken.
Introduction
Oligodendrocytenafstammingscellen - inclusief oligodendrocytenvoorlopercellen (OPC's), myeliniserende oligodendrocyten en overgangstypen daartussenin - vormen een belangrijke groep menselijke hersencellen1 die actief deelnemen aan vele kritieke functies voor de juiste werking en het onderhoud van ons centrale zenuwstelsel tijdens de neurale ontwikkeling en veroudering 2,3,4 . Hoewel oligodendrocyten bekend staan om het produceren van myeline om de overdracht van neuronale activiteit te vergemakkelijken en de axonale gezondheid in witte stof te ondersteunen, zijn OPC's overvloedig (~ 5%) in grijze stof waar myelinisatie schaars is en voeren activiteitsafhankelijke signaleringsfuncties uit om leergedrag en geheugenvorming te regelen 5,6,7,8 . Hoe oligodendrogliale cellen functioneren en disfunctie in de pathogenese van de ziekte van Alzheimer (AD) en andere leeftijdsgebonden neurodegeneratieve aandoeningen is onderbelicht9. De tekortkomingen van een geschikt modelsysteem en tekortkomingen in de algemene kennis om een experimenteel pad voorwaarts te begeleiden, zijn de belangrijkste redenen voor deze kloof.
In het licht van de laatste doorbraken in het afleiden van menselijke hersencellen uit pluripotente stamcellen, waaronder embryonale stamcellen (ES) en geïnduceerde pluripotente stamcellen (iPS), zijn dergelijke cellulaire modellen in combinatie met moderne genbewerkingstools naar voren gekomen als robuuste hulpmiddelen om de ingewikkelde nexus van cellulaire interacties in de hersenen aan te pakken, en zijn ze in staat om mensspecifieke ziekteverschijnselen aan te tonen10, 11. Gezien het feit dat individuele hersenceltypen verschillende en zelfs conflicterende effecten kunnen vertonen in het licht van dezelfde AD-bevorderende aandoeningen12,13, biedt deze stamcelmethodologie op unieke wijze celtypespecifieke informatie die eerder is gemist met behulp van gevestigde in vivo of in vitro modellen die alleen geaggregeerde uitlezingen bieden van verzamelingen hersenceltypen. In het afgelopen decennium zijn een groot aantal betrouwbare protocollen ontwikkeld om menselijke neuronen te genereren uit transdifferentiatie van ES / iPS-cellen of directe conversie van andere terminaal gedifferentieerde celtypen (bijv. Fibroblasten)14,15. In het bijzonder kan de toepassing van belangrijke neurogene transcriptiefactoren (bijv. neurogenine 2, Ngn2)16 op menselijke pluripotente stamcellen een homogene populatie van goed gekarakteriseerde neuronale celtypen genereren voor zuivere culturen zonder dat er behoefte is aan coculturing met gliacellen 12,17,18. Voor geïnduceerde menselijke oligodendrocyten zijn er een paar gepubliceerde protocollen die functionele cellen kunnen genereren die sterk lijken op hun primaire tegenhangers, met een breed scala aan efficiëntie en vraag in tijd en middelen 19,20,21,22,23,24,25,26,27,28 . Tot op heden is geen van deze protocollen toegepast om te onderzoeken hoe oligodendrogliale cellen reageren op en invloed hebben op AD-pathogenese.
Hier beschrijven we onze verbeterde protocollen voor enkelvoudige en gemengde culturen van door de mens geïnduceerde neuronen (iN's) en OPC's/oligodendrocyten (iOPCs/iOL's). Het hier beschreven iN-protocol is gebaseerd op de veelgebruikte Ngn2-benadering16 en heeft als extra eigenschap gliavrij te zijn. De resulterende iNs zijn homogeen en lijken sterk op de corticale laag 2/3 exciterende neuronen, met karakteristieke piramidale morfologie, genexpressiepatroon en elektrofysiologische kenmerken17,18 (figuur 1). Om enkele van de fundamentele barrières bij gerichte differentiatie van pluripotente stamcellen te overwinnen, hebben we een eenvoudige en effectieve methode ontwikkeld voor de voorbehandeling van lage doses dimethylsulfoxide (DMSO)29,30, en rapporteerden we een verhoogde neiging van menselijke ES / iPS-cellen om te transdifferentiëren in iOPCs en iOL's31, gebaseerd op een breed aangepast protocol van Douvaras en Fossati32 . We hebben het protocol verder vereenvoudigd en een robuuste differentiatiebevorderende verbinding, clemastine 7,33,34, opgenomen om het proces van oligodendrogliale rijping te versnellen. Als gevolg hiervan (figuur 2) kunnen de iOPCs worden gegenereerd in 2 weken (~ 95% positief voor de marker O4) en iOL's in vier weken (met volwassen markers MBP en PLP1). Interessant is dat we ontdekten dat iOPCs alleen een opmerkelijke hoeveelheid amyloïde-β (Aβ) afscheiden, consistent met de onafhankelijke transcriptomische gegevens die de overvloedige expressie van het amyloïde precursoreiwit (APP) en het verwerkingsprotease β-secretase (BACE1) in oligodendrocytenlijncellen35,36 aantonen. Bovendien bevordert ons iN-iOPC cocultuursysteem de opwarming van axonen door MBP-positieve iOL-processen en biedt het aanzienlijke ondersteuning voor synapsvorming (figuur 3). De protocollen die we hieronder hebben beschreven, hebben dus technische en biologische voordelen ten opzichte van eerder gecatalogiseerde neuron-oligodendroglia co-cultiveringsmethoden en houden een belofte in bij het beter modelleren van de neurodegeneratie in AD.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Inductie van menselijke neuronen uit menselijke pluripotente stamcellen
- Lentivirus voorbereiding (~ 5 dagen, gedetailleerd protocol zoals eerder beschreven16)
- Plaat ~ 1 miljoen HEK293T-cellen per T75-kolf, om ze ~ 40% confluent te hebben bij het uitvoeren van transfectie. Transfecteer ze met plasmiden die tetracycline-induceerbare Ngn2 en puromycine-resistent gen (PuroR; onder dezelfde TetO-promotorcontrole) tot expressie brengen, rtTA en de drie helperplasmiden pRSV-REV, pMDLg / pRRE en VSV-G (12 μg lentiviral vector-DNA en 6 μg van elk van het helperplasmide-DNA). Bereid ten minste drie kolven per lentiviruspreparaat. Gebruik PEI voor transfectie volgens de instructies van de fabrikant. Vervang de media na 16 uur en gooi deze weg.
- Oogst vrijgekomen virale deeltjes door elke dag kweekmedia te verzamelen en gedurende 3 dagen te vervangen door verse media. Pool de verzamelde media met virale deeltjes voor zuivering. Filter het virus door een filter van 0,22 μm en centrifugeer gedurende 90 minuten op 49.000 x g . Resuspendeer de pellet in het juiste volume PBS-glucose (~150 μL).
- Neuron Inductie (~5 dagen)
OPMERKING: Dit inductieprotocol (figuur 1A; stroomdiagram) is zeer effectief voor zowel iPS- als ES-cellen met gevalideerde pluripotentie (die kan worden getest door immunohistochemische kleuring van goed gekarakteriseerde pluripotentiemarkers; Figuur 1B).- Gebruik in de handel verkrijgbare H1 menselijke ES-cellen bij het passeren van 52 (zie Materiaaltabel). Kweek de cellen op extracellulaire matrixoplossing gecoate 6-well platen (~ 0,5 mg matrixoplossing per 6-well plaat; zie Tabel van materialen) met behulp van ES-celonderhoudsmedium (zie Tabel van materialen) en incubeer de platen bij 37 °C met 5% CO2.
- Maak op dag -2 ES-cellen (80% confluent) los met 1 ml celloslatingsoplossing (zie Materiaaltabel) en incubeer bij kamertemperatuur gedurende 10 minuten. Breng de cellen over naar een buis; was de put met 2 ml media en combineer in dezelfde buis. Centrifugeer bij 300 x g gedurende 5 minuten, resuspenseer de pellet in media en plaats de cellen op matrix gecoate 6-well platen met de zaaidichtheid van 1 x 105 cellen per put.
- Voeg op dag -1 lentivirussen die Ngn2 plus PuroR en rtTA tot expressie brengen samen met polybreen (8 μg/ml) toe aan de ES-cellen in vers ES-celonderhoudsmedium (zie materiaaltabel). De exacte hoeveelheid virussen moet worden bepaald door de werkelijke titers of de titratie. We voegen meestal 5 μL per virus per put toe in een 6-well plaat.
- Voeg op dag 0 Doxycycline (2 μg / ml, om Ngn2-expressie te activeren) toe in DMEM-F12-medium met N2-supplement zonder morfogenen.
- Voeg op dag 1 Puromycine toe in vers medium DMEM-F12 plus N2 en doxycycline, tot de uiteindelijke concentratie van 1 μg / ml medium. Selecteer de getransduceerde cellen in Puromycine gedurende ten minste 24 uur. Een hogere puromycineconcentratie (tot 5 μg / ml) en een langere selectieperiode (tot 48 uur) kunnen nodig zijn om de ondergetransduceerde cellen adequaat te verwijderen als de virustiter laag is.
- Maak op dag 2 differentiërende neuronen los met celloslatingsoplossing (zie Tabel met materialen) en plaats ze opnieuw op 24-putplaten (tussen 80.000-200.000 cellen / put) bedekt met matrixoplossing (zie Materiaaltabel) en onderhoud ze in NBA / B27-medium zonder doxycycline. De zaaidichtheid is van cruciaal belang.
- In dit stadium kunnen losgemaakte neuronen worden ingevroren in een gespecialiseerd commercieel vriesmedium (zie Materiaaltabel) en tot 3 maanden worden opgeslagen in vloeibare stikstof. Zuivere neuronen kunnen worden verguld, goed voor de typische ~ 15% -20% celdood na dooi, alleen gekweekt of samen gekweekt met andere hersenceltypen (zie stap 3.2.3. voor co-kweken met OPC's).
- Kweek zuivere iNs op de platen die zijn gecoat met extracellulaire matrixgebaseerde oplossingen volgens de instructies van de fabrikant (zie materiaaltabel). De karakteristieke piramidale morfologie moet duidelijk zijn op dag 4 (en dag 6; Figuur 1C). De synapsvorming kan al op dag 14 tot 16 worden gedetecteerd en is prominent aanwezig op dag 24 door immunohistochemische kleuring met standaard pre- en post-synaptische markers. (Figuur 1D; gelabeld met de pre-synaptische marker Synapsin 1 en de dendritische marker Map2).
2. Inductie van humane oligodendrocytenprecursorencellen (OPC's) uit pluripotente stamcellen en rijping van oligodendrocyten
- Neural Progenitor Cell (NPC) generatie: monolayer protocol (~7 dagen). Zie figuur 2A voor het stroomdiagram.
- Kweek H1 menselijke ES-cellen zoals eerder beschreven (zie stap 1.2.1.) en trans-differentieer ze in neurale voorlopercellen (NPC's) door een gevestigde aanpak genaamd duale SMADi, met kleine molecuulremmers voor meerdere signaalroutes. Hier gebruiken we een algemeen aanvaarde commerciële kit en volgen we het monolayer-protocol van de fabrikant (zie Materiaaltabel).
- Op dag -1, plaat 0,5-1 x 106 cellen per put in een 6-well plaat gecoat door een groeifactor gereduceerde matrixoplossing (zie Tabel van materialen; ~ 0,5 mg matrixoplossing per 6-well plaat) met ES-cel onderhoudsmedium (zie Tabel van materialen). Deze groeifactor gereduceerde matrixoplossing wordt gebruikt om alle platen te coaten die in de volgende stappen zullen worden gebruikt.
- Behandel cellen op dag 0 gedurende 24 uur met het onderhoudsmedium van de ES-cel (zie materiaaltabel) aangevuld met 2% DMSO.
- Verander op dag 1-6 de volledige media met warm (37 °C) neuraal inductiemedium dat de SMAD-remmers uit de commerciële kit bevat (zie Materiaaltabel). Als cellen zich vóór dag 7 delen en samenvloeien, breng ze dan door naar de zaaidichtheid van 0,5-1 x 106, zoals eerder beschreven in stap 2.1.2.
- Op dag 7, passage NPC's met behulp van celloslatingsoplossing (zie tabel met materialen) en plaat met een zaaidichtheid van 1-2 x 105 cellen / put van een 24-well plaat.
- Test de differentiatie-efficiëntie door immunohistochemische (IHC) kleuring op afwezigheid van pluripotentiemarker, OCT4 bijvoorbeeld, en aanwezigheid van NPC-markers zoals PAX6, Nestin en Sox1.
- In dit stadium kunnen losse NPC's worden ingevroren in de gespecialiseerde commerciële NPC-vriesmedia (zie Materiaaltabel) en tot 3 maanden worden opgeslagen in vloeibare stikstof. Na één keer bevriezen en dooien behouden NPC's nog steeds de multipotentie om neuronen, astrocyten en OPC's met betrouwbare protocollen te laten ontstaan.
- Oligodendrocyte precursor cell (OPC) generatie (~ 7 dagen). Zie figuur 2A voor het stroomdiagram.
- Op dag 7, passeer NPC's met behulp van celloslatingsoplossing (zie Tabel van materialen) en plaats ze met een zaaidichtheid van 1-2 x 105 cellen per put in een 24-well plaat in warm (37 °C) neuraal inductiemedium plus SMAD-remmers uit de commerciële kit (zie Tabel van materialen).
- Bereid op dag 8 een oplossing van 1% DMSO in het OPC-differentiatiemedium en behandel de vergulde NPC's gedurende 24 uur. Het OPC-differentiatiemedium bestaat uit: DMEM/F12-medium, 1% N2-supplement, 1% B27-supplement, bFGF bij 20 ng/ml, SAG bij 1 μM, PDGF-AA bij 10 ng/ml (zie materiaaltabel).
- Vervang op dag 9 media door een nieuw OPC-differentiatiemedium zonder DMSO. Voer de cellen om de dag tot dag 15. Als de cellen vóór dag 15 samenvloeiing bereiken, breng ze dan door naar de zaaidichtheid van 1-2 x 105 cellen per put zoals beschreven in stap 2.2.1.
- Op dag 14, plaat-OPC's in OPC-differentiatiemedium met een dichtheid van 1-2 x 105 cellen / put in een 24-well plaat.
- Test in dit stadium (dag 15) cellen op de aanwezigheid van OPC-specifieke markers door IHC-kleuring of qPCR (bijv. O4, Olig1/2, CSPG4/Ng2, NKX2.2, PDGFRa; Figuur 2B) en voor de afwezigheid van NPC-markers (Pax6 of Nestin; Figuur 2D). We detecteren meestal de O4-immunoreactiviteit in meer dan 95% van de cellen op dag 15. Van bijzonder belang voor de ziekte van Alzheimer is de expressie van APP (amyloïde precursoreiwit), BACE1 (het verwerkingsprotease β-secreatase 1) en peptide-amyloïde-β (Aβ) overvloedig aanwezig in OPC's (figuur 2F).
- Oligodendrocyten (OL) rijping (~7-20 dagen)
- Vervang op dag 15 media door OL-rijpingsmedium: Neurobasal-A-medium, 2% B27-supplement, 1 μM cAMP, 200 ng / ml T3 triiodothyronine en Clemastine van 1 μM (zie materiaaltabel). Verander het medium om de dag of elke dag, indien nodig.
- Wanneer cellen 90% confluentie bereiken, splits je ze in een verhouding van 1:3 tot 2 passages of totdat de celdeling aanzienlijk vertraagt. Als OPC's te snel delen en in minder dan 3 dagen samenvloeiing bereiken, voeg dan Ara-C (zie Materiaaltabel) toe in een concentratie van 2-5 μM gedurende 1-3 dagen. Actieve proliferatie duidt op een verlaagde rijpingsefficiëntie.
- Onderzoek de efficiëntie van oligodendrogliale rijping door de expressie van OL-markers te beoordelen, bijvoorbeeld CLDN11, PLP1, MBP door qPCR, IHC-kleuring of immunoblotting. De karakteristieke morfologie van zeer complexe structuren (figuur 2C) en de expressie van OL-markers (figuur 2E) moeten gemakkelijk worden gedetecteerd op dag 28.
3. Co-kweken van door de mens geïnduceerde neuronen (iNs) en oligodendrocytenvoorlopercellen (iOPCs)
- iOPC-beplating (~ 3 dagen)
- Plaat-iOPCs op dag 14 met een dichtheid van 1 x 105 cellen per put in een 24-well plaat (zoals hierboven beschreven in stap 2.2.4.) in OPC-differentiatiemedium (zoals beschreven in stap 2.2.2.).
- iN-iOPC co-cultuur opgezet
- Maak op dag 15 de geïnduceerde menselijke neuronen los in de stap van dag 2 na de puromycineselectie (zoals beschreven in stap 1.2.6.) met celloslatingsoplossing (zie materiaaltabel).
- Voeg neuronen toe aan de gekweekte OPC's, plating met de zaaidichtheid van 2 x 105 cellen per put in de 24-well plaat met groeiende OPC's (vanaf stap 3.1.1). Gebruik het co-kweekmedium met Neurobasal-A medium, 2% B27 supplement en 100 ng/ml T3 triiodothyronine. Verander het medium op de volgende dag en daarna om de andere dag. Als OPC's zich te snel vermenigvuldigen en in minder dan 3 dagen samenvloeiing bereiken, voeg dan Ara-C toe in een concentratie van 2-5 μM. Een representatief beeld van de iN's en iOPC's die na 7 dagen in cocultuur zijn gegroeid, wordt weergegeven in figuur 3A.
- Gebruik bevroren neuronen die zijn bereid zoals hierboven beschreven in stap 1.2.7 voor co-kweek met OPC's. Plaat bevriezen-en-ontdooien neuronen met een hogere dichtheid van 3 x 105 cellen per put.
- Na dag 14-16 in co-culturen kan de synapsvorming in iNs worden waargenomen door IHC-kleuring van pre- en post-synaptische markers, en op dag 21 zou de synaptische puncta overvloedig moeten zijn (figuur 3C) en neuronale activiteiten kunnen betrouwbaar worden geregistreerd.
- Vanaf dag 21 test u cellen voor OL-specifieke markers (bijvoorbeeld MBP en PLP1). Op dag 28 observeren we normaal gesproken het fenomeen ensheathing van iN-axonen door iOL-processen, gelabeld door IHC-kleuring voor specifieke markers (figuur 3B; neurofilament NF voor iN-axonen en MBP voor iOPC-processen).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Directe generatie van door de mens geïnduceerde neuronen uit menselijke pluripotente stamcellen
Het is van groot belang dat de startende menselijke pluripotente stamcellen een hoge mate van pluripotentie vertonen voor een succesvolle generatie van iN's of iOPCs/iOL's. Daarom moeten cellen worden gekleurd voor specifieke markers, zoals Oct4 en SOX2, voordat ze beginnen met een van de inductieprotocollen die in dit manuscript worden beschreven (figuur 1A). Menselijke H1-cellen werden gebruikt om geïnduceerde exciterende voorhersenenneuronen te verkrijgen volgens het eerder gepubliceerde protocol door Zhang et al. met enkele wijzigingen (figuur 1C)12,16,17,18. Hier presenteren we een protocol waarin iN's op dag 2 opnieuw worden verguld in pure cultuur op matrixoplossing (zie tabel met materialen), bij afwezigheid van een feederlaag: glia of fibroblasten. Naast de eerder gepubliceerde protocollen, merken we op dat het bevriezen van iNs op dag 2 geen significante invloed heeft op de levensvatbaarheid van cellen (~ 15% -20% celdood na ontdooien). Zuivere neuronen in cultuur zullen synapsine1 gaan uitdrukken op dag 14-16 (figuur 1D). Het vaststellen van een zuivere neuronale cultuur is erg belangrijk omdat bepaalde factoren, bijvoorbeeld de leidende AD-risicofactor ApoE, tot expressie kunnen worden gebracht door cellen in de feederlaag en dit kan de resultaten aanzienlijk verstoren.
iOPC-generatie en iOL-rijping wordt verbeterd door DMSO-behandeling
Hier presenteren we een snel en efficiënt protocol dat het genereren van iOPCs in 2 weken en volwassen iOL's in 4-5 weken mogelijk maakt (figuur 2A). We maakten gebruik van de methode van voorbijgaande DMSO-behandeling die we eerder ontwikkelden om de differentiatie-efficiëntie voor ES- en iPS-cellen te vergroten 29,30,31. DMSO-behandeling verrijkt het aantal cellen in de vroege G1-fase voor een betere signaleringsintegratie, wat differentiatie bevordert. We voerden de eerste behandeling uit voordat we menselijke ES-cellen induceerden om NPC's te genereren, en de tweede behandeling voordat we NPC's differentieerden in iOPCs. We kunnen specifieke OPC-markers (Olig2, CSPG4, NKX2.2 en PDGFRA) al 2 weken na plating van ES H1-cellen detecteren (figuur 2B, E). De iOPC-populatie in dit stadium is vrij homogeen, met >95% van de cellen positief voor O4-kleuring en een hoog niveau van immunoreactiviteit voor andere markers (figuur 2B). Na het begin van de OL-rijping op dag 15 kunnen we meestal specifieke OL-markers (MBP, O1, CLDN11 en PLP1) detecteren vanaf dag 28 (figuur 2C, E). De expressie van deze stadiumspecifieke markers correleert met het ontwikkelingsverloop van oligodendrogliale cellen en suggereert een versneld tempo, waarbij de NPC-markers geleidelijk dalen, OPC-markers pieken rond de tweede week en OL-markers stijgen tegen de derde week (figuur 2D, E) 37. Houd er rekening mee dat dit rijpingsproces de celpopulaties diversifieert. De subpopulaties in continuüm, bestaande uit meerdere tussenstadia tussen OPC's en volwassen myeliniserende oligodendrocyten, kunnen aanwezig zijn en verantwoordelijk zijn voor een wisselend percentage van de totale cellen, waarbij meer volwassen cellen op een later tijdstip domineren.
Ter vergelijking: we kochten de iOC's met veel referenties en rijpten ze in iOL volgens het protocol van de fabrikant. We hebben de expressie van de hierboven genoemde markers getest in zowel onze iOPC- als iOL-preparaten en in de cellen die we hebben gekocht. We stelden vast dat de cellen die volgens ons protocol werden gegenereerd, een hogere expressie hadden van alle geteste genen (figuur 2E). Interessant is dat toen we de uitgescheiden niveaus van twee belangrijke isovormen van amyloïde-β (Aβ40 en Aβ42) in iN's versus iOPC's testten, we merkten dat iOPCs meer van beide fragmenten uitscheidden, maar de verhouding bleef hetzelfde (figuur 2F).
Co-culturing van iN's en iOPC's
Dit protocol is speciaal geoptimaliseerd voor het co-kweken van iN's en iOPC's en maakt onze real-time monitoring van de intercellulaire communicatie tussen deze twee celtypen in de loop van de neurale ontwikkeling mogelijk. De ideale platingdichtheden voor beide celtypen moeten worden bepaald met een reeks celnummertitratie om een goede differentiatie te bereiken (figuur 3A). Na 4 weken in coculturen wordt verwacht dat de iOPC's voldoende worden gedifferentieerd in OK's die positief zijn voor specifieke markers zoals MBP en processen uitbreiden naar ensheath axonen (figuur 3B). Het co-kweeksysteem kan het aantal synapsen sterk verhogen, wat aangeeft dat de iOPC's een neuronale ondersteuning bieden door fysieke contacten of het vrijgeven van trofische factoren (figuur 3C). We kunnen de co-culturen tot 6 weken in aanvaardbare gezondheidstoestand houden en vaststellen dat het synapsgetal en andere neuronale kenmerken rond de vijfde week plateau. Van belang is dat astrocyten en microglia niet aanwezig zijn in onze preparaten en hun afwezigheid kan worden gedocumenteerd door de expressie van specifieke markers te controleren (figuur 3D). De iOPCs drukken een groot aantal goed gekarakteriseerde genen uit die mogelijk kunnen reageren op en bemiddelen op de activiteitsafhankelijke signalen van naburige neuronen, in een paracriene (bijv. neurotrofines en metabolieten) en / of op een synaptische manier (figuur 3E en 3F).

Figuur 1: Directe generatie van door mensen geïnduceerde neuronen (iN's) uit hPSC's. (A) Stroomdiagram van iN-generatie. (B) Representatieve heldere veld- en immunofluorescentiebeelden van de uitgangscultuur van menselijke pluripotente stamcellen (H1) om de pluripotentie te bevestigen. Oct4 wordt weergegeven in rood en Sox2 in groen. (C) Representatieve heldere veldbeelden van iNs op dag 4 en dag 6. (D) De karakteristieke morfologie voor dendritische arborisatie en synaps puncta in iNs gekweekt in zuivere cultuur gedurende 24 dagen en gekleurd door immunofluorescentiekleuring voor dendritische marker Map2 en pre-synaptische marker Synapsin 1 (Syn1). Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 2: iOPC-generatie en iOL-rijping. (A) Stroomdiagram van iOPC- en iOL-generatie. (B) Representatieve beelden van helder veld en immunofluorescentie van iOPCs op dag 15. Olig2 (pan-oligodendroglia marker) wordt weergegeven in groen, O4 (OPC marker) in rood en DAPI in blauw. Uit de beeldvorming bleek dat >95% van de iOPC's positief is voor O4 en 25% voor Olig2. (C) Representatieve beelden van helder veld en immunofluorescentie van iOL's op dag 28. MBP wordt weergegeven in groen, O1 in rood en DAPI in blauw. (D) De expressie van NPC-marker PAX6 neemt dramatisch af in iOPCs op dag 14 en daalt verder naar achtergrond in OK's op dag 28, wat wijst op een robuuste NPC-transdifferentiatie en een hoge mate van homogeniteit in de iOPC-populatie. (E) Het tijdsverloopexpressieprofiel van gemeenschappelijke OPC- en OL-markergenen in culturen gegenereerd door het beschreven protocol, zonder (-DMSO) of met (+DMSO) de stap van DMSO-incubatie (stappen 2.1.3 en 2.2.2), getest op verschillende tijdstippen. Ter vergelijking: commerciële iOPC's (zie Materiaaltabel) werden gerijpt volgens de instructies van de fabrikant en zowel iOPC's (iOPC-Tempo) als iOL's (iOL-Tempo) werden getest op dezelfde markers. Zoals verwacht werd MBP (een volwassen oligodendrocytmarker) niet gedetecteerd (N.D.) in de vroege stadia van differentiatie in alle geteste iOPC's. De DMSO verbeterde de efficiëntie van OPC-differentiatie en OL-rijping aanzienlijk. (F) De productie en secretie van Aβ40 en Aβ42 in zuivere RN's en iOPC-culturen, gemeten met commerciële ELISA-kits (zie Materiaaltabel) op supernatant verkregen uit zuivere iN's en iOPC-culturen, zowel op dag 15 als genormaliseerd door celaantallen (beide met de dichtheid van 200.000 cellen per put in een plaat met 24 putten). Klik hier om een grotere versie van deze figuur te bekijken.
Gegevens in staafdiagrammen worden uitgezet als gemiddelde ± SEM (n ≥ 3). Statistische significantie werd geëvalueerd door student t-test (*, p < 0,05; ***, p < 0,001); in (D), vergeleken met de NPC; in (E), vergeleken met de regeling iOPC-Tempo; in (F), vergeleken met iN.

Figuur 3: Cocultuur van iN's en iOPC's. (A) Representatief helder veldbeeld van co-gekweekte iN's en iOPC's op dag 7, met een juiste dichtheid voor verdere rijping. (B) Representatief immunofluorescentiebeeld van iN's en iOPC's dat gedurende 28 dagen samen is gekweekt. Axonale marker neurofilament NF wordt weergegeven in groene en oligodendrocytische marker MBP in rood. Rechts, een segment van iN axon verwarmd door iOL-proces (MBP +). (C) Synapsvorming getest in 4 weken oude coculturen. Cellen werden gekleurd voor Synapsin 1 (Syn1, groen) en MAP2 (rood), en synaptische puncta werden gekwantificeerd door confocale analyse van de dichtheid langs de dendritische segmenten zoals beschreven17,18. (D) In onze coculturen van iN's en iOPC's (7 dagen co-kweken) is de expressie van astrocytenmarkers, ALDHL1 en GFAP, minimaal (boven) en wordt de expressie van microgliamarkers, TMEM119, TREM2 en CD33, niet gedetecteerd (N.D.) door qPCR. De besmetting van deze twee gliaceltypen is dus uitgesloten. (E) Het samentellen van iOPC met iN leidt tot de vorming van neuron-OPC synapsen. De fluorescentie-gelabelde post-synaptische marker PSD95-mCherry wordt alleen uitgedrukt in OPC's en vertoont een diffuus patroon in afzonderlijke culturen (links) maar aggregaat om puncta te vormen in coculturen (rechts, aangegeven met pijlen; Tuj1, neuronale marker). (F) De expressie van goed gekarakteriseerde oligodendrogliale genen die neuronale activiteiten in de zuivere culturen van iOPCs op dag 14 kunnen waarnemen en erop kunnen reageren. Klik hier om een grotere versie van deze figuur te bekijken.
Gegevens in staafdiagrammen worden uitgezet als gemiddelde ± SEM (n ≥ 3). Statistische significantie werd geëvalueerd door student t-test (**, p < 0,005; ***, p < 0,001); in (C), vergeleken met de voorwaarde geen OPC; in (D), vergeleken met primaire astrocyten in het bovenste paneel.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Naast de fysieke en metabole ondersteuning om de synapsstructuren te stabiliseren en de zoute signaalgeleiding door myelinisatie te vergemakkelijken, kunnen oligodendrocytenafstammingscellen een neuronaal activiteitspatroon vormen via snelle en dynamische kruisgesprekken met neuronen 5,6,7. Terwijl in de AD-pathologie de oligodendrogliale responsen aanvankelijk werden beschouwd als slechts secundair aan ontsteking en oxidatieve stress, is er nu veelbelovend bewijs dat de gecompromitteerde myeline-integriteit een vroege pathogene gebeurtenis is voorafgaand aan het verschijnen van Aβ-aggregatie en tau-hyperfosforylering9. Bovendien is het herstel van myelinisatie door zelfvernieuwing van OPC's bijzonder kwetsbaar in AD38, een proces dat sterk afhankelijk is van neuronale activiteiten. Het begrijpen van het mechanisme ter ondersteuning van gezonde neuron-oligodendrocyten signalering is dus een uitstekende gelegenheid voor het identificeren van nieuwe therapeutische doelen.
Het Ngn2-protocol met enkele transcriptiefactor is een van de meest gebruikte technieken voor het genereren van van stamcellen afgeleide menselijke neuronen, en de hier beschreven procedures zijn verdere verfijningen voor het verkrijgen van zuivere neuronale culturen. Ons iOPC/iOL-protocol heeft een inductieperiode die korter is dan de eerder gepubliceerde studies (4 tot 24 weken), met een robuuste opbrengst en zuiverheid vergelijkbaar met andere veelgebruikte protocollen 19,20,22,23,24,25,26,27,28 . Ons protocol introduceert de stapsgewijze differentiatie van ES-cellen naar NPC's, OPC's en ten slotte oligodendrocyten door gekarakteriseerde patroonsignalen en genereert functionele cellen die kunnen worden gebruikt om de regulatie van myelinisatiehomeostase te bestuderen en in vitro of in vivo te repareren (bijvoorbeeld door engraftment in het rillermuismodel) zoals beschreven in het vorige werk. De verbetering van ons protocol wordt sterk bevorderd door de DMSO-incubatie, die het retinoblastoomeiwit activeert en de G1-fase van de celcyclus verlengt om de stimuli van gerichte differentiatie beter te integreren, en ook de terminale differentiatie in functionele derivaten verbetert29,30. Ten slotte verkort het gebruik van clemastine, een muscarine- en antihistaminische verbinding die wordt geïdentificeerd door geneesmiddelscreening voor remyelinisatietherapieën33, bovendien de rijping van oligodendrocyten, zoals waargenomen bij iPS-celpreparatie en levende dieren 7,28.
De beperkingen van de techniek liggen vooral in de intrinsieke discrepantie tussen de vereenvoudigde in vitro settings en de in vivo omgevingen in de hersenen; deze discrepantie leidt tot een vermindering van het volledige ontwikkelingspotentieel in de gevorderde stadia voor individuele hersenceltypen. Voor iNs waren recente studies in staat om de culturen gedurende een aanzienlijk lange periode in goede synaptische gezondheid te houden, maar onthulden nog steeds enige relatieve onvolwassenheid die zich manifesteerde als verminderde wervelkolomachtige structuren en verminderde spontane synaptische transmissie in "oude" iN-culturen (zelfs de 25 maanden oude)39. Hoewel reproduceerbaar is aangetoond dat iOPC's axonen in vivo myeliniseren nadat ze in transgene muizenhersenen zijn getransplanteerd, vormen de in vitro myelinisatietests met elektronenmicroscopische evaluatie nog steeds een technische uitdaging met onbevredigende efficiëntie voor bijna alle gepubliceerde protocollen19,28, inclusief deze. Daarom wordt niet verwacht dat ons neuron-OPC co-kweeksysteem het hersenverouderingsproces en het late stadium van AD-pathologie getrouw zal nabootsen. Integendeel, het is uniek klaar om de uitgebreide intercellulaire interacties tussen neuronen en OPC's of oligodendrocyten in een vroeg stadium te ontwarren, die onafhankelijk zijn van myelinisatie en toch fundamenteel zijn voor een goede neurale ontwikkeling en ziektepathogenese.
Elk van de drie hier beschreven procedures heeft zijn belangrijkste stappen binnen het protocol en kan wijzigingen en probleemoplossing vereisen. Voor het iN-protocol zijn er twee kritieke stappen: puromycineselectie (stap 1.2.5) en platingdichtheid (stap 1.2.6). Onvolledige verwijdering van ondergetransduceerde cellen resulteert in de besmetting van slecht gedifferentieerde cellen en brengt de neuronale overleving en functies in gevaar. De modificaties voor een sterkere puromycineselectie met een hogere concentratie en langere incubatie zoals beschreven in stap 1.2.5 zouden moeten worden overwogen. De geschikte platingdichtheid moet worden bepaald door titratie voor elke pluripotente cellijn, omdat een lage dichtheid leidt tot instorting van de culturen en een hoge dichtheid de celaggregatie bevordert en de neuronale groei belemmert. Voor het iOPC/iOL-protocol zijn de twee kritieke stappen de controle van celproliferatie in OPC-differentiatie (stap 2.2.3) en platingdichtheid voor OL-rijping (stappen 2.2.4 en 2.3.2). De overgroei van differentiërende NPC's signaleert een slechte respons op de OPC-differentiatieprikkels en moet worden gedempt door een geschikte dosering ara-c-behandeling (binnen het aangegeven bereik). Terwijl opc's voor rijping worden plat, heeft hier de voorkeur aan een lager bereik van celdichtheid, omdat de schaarse verdeling de inductie van een fysiologische morfologie van complexe structuren kan vergemakkelijken (zoals weergegeven in figuur 2C). Voor het iN-iOPC co-kweekprotocol willen we de aandacht vestigen op de kritieke stap van plating met een geschikte dichtheid voor beide celtypen (stappen 3.1.1 en 3.2.2). In het bijzonder hechten de iN's zich mogelijk niet goed aan het oppervlak tussen groeiende OPC's en hebben ze de neiging om eerst los te komen wanneer de cultuur samenvloeiing bereikt. De optimale verhouding zou moeten worden bepaald door de celnummers te titreren.
Over het algemeen is deze reductionistische benadering die in onze protocollen zit een krachtig hulpmiddel om de specifieke hetero-cellulaire interacties te ontleden van de inherente complexiteit van het menselijk brein, en dienen om oligodendrogliale biologie in gezondheid en in AD te ontdekken. De betekenis ten opzichte van bestaande methoden is naar onze mening dus vrij duidelijk. Een bijkomend nut van de hier ontwikkelde methoden voor de toekomstige toepassingen is celgebaseerde therapie voor demyeliniserende aandoeningen, zoals post-radiotherapie40 en dwarslaesie 41,42. Bovendien kan de hoge doorvoercapaciteit van dit op stamcellen gebaseerde systeem ook op grotere schaal worden gebruikt om bibliotheken van kleine moleculen te screenen op verbindingen die de fysiologische status van neuronen, OPC's, oligodendrocyten en hun interacties kunnen beschermen of herstellen. Daarom geloven we dat de hier beschreven protocollen toekomstig werk zullen vergemakkelijken bij het ontwikkelen van betere modelleringstools en effectieve behandelingen voor AD en andere neurodegeneratieve aandoeningen.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs hebben niets te onthullen.
Acknowledgments
Dit werk werd ondersteund door de subsidies van National Institutes of Health (R00 AG054616 aan Y.A.H. en T32 GM136566 aan K.C.), Stanford University School of Medicine en een Siebel Fellowship (toegekend aan S.C.). Y.A.H. is een GFL Translational Professor van het Center for Translational Neuroscience in het Brown Institute for Translational Sciences.
Materials
| Name | Company | Catalog Number | Comments |
| Accutase | STEMCELL Technologies | 7920 | |
| B27 supplement | ThermoFisher | 17504044 | |
| bFGF | ThermoFisher | PHG 0266 | |
| cAMP | MilliporeSigma | A9501 | |
| Clemastine | MilliporeSigma | SML0445 | |
| DMEM/F12 medium | STEMCELL Technologies | 36254 | |
| DMSO | ThermoFisher | D12345 | |
| Doxycycline | MilliporeSigma | D3072 | |
| Fetal Bovine Serum | ScienCell | 10 | |
| H1 human ES cells | WiCell | WA01 | |
| Matrigel | Corning | 354234 | |
| mTeSR plus | STEMCELL Technologies | 5825 | |
| N2 supplement | ThermoFisher | 17502001 | |
| Neurobasal A medium | ThermoFisher | 10888-022 | |
| Non Essential Amino Acids | ThermoFisher | 11140-050 | |
| PDGF-AA | R&D Systems | 221-AA-010 | |
| PEI | VWR | 71002-812 | |
| pMDLg/pRRE | Addgene | 12251 | |
| Polybrene | MilliporeSigma | TR-1003-G | |
| pRSV-REV | Addgene | 12253 | |
| Puromycin | ThermoFisher | A1113803 | |
| ROCK Inhibitor Y-27632 | STEMCELL Technologies | 72302 | |
| SAG | Tocris | 4366 | |
| STEMdiff Neural Progenitor Freezing Media | STEMCELL Technologies | 5838 | |
| STEMdiff SMADi Neural Induction Kit | STEMCELL Technologies | 8581 | |
| T3 triiodothyronine | MilliporeSigma | T6397 | |
| Tempo-iOlogo: Human iPSC-derived OPCs | Tempo BioScience | SKU102 | |
| TetO-Ng2-Puro | Addgene | 52047 | |
| VSV-G | Addgene | 12259 |
References
- Pelvig, D. P., Pakkenberg, H., Stark, A. K., Pakkenberg, B. Neocortical glial cell numbers in human brains. Neurobiology of Aging. 29 (11), 1754-1762 (2008).
- Barres, B. A. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 60 (3), 430-440 (2008).
- De Strooper, B., Karran, E. The cellular phase of Alzheimer's disease. Cell. 164 (4), 603-615 (2016).
- Monje, M. Myelin plasticity and nervous system function. Annual Review of Neuroscience. 41, 61-76 (2018).
- Hughes, E. G., Orthmann-Murphy, J. L., Langseth, A. J., Bergles, D. E. Myelin remodeling through experience-dependent oligodendrogenesis in the adult somatosensory cortex. Nature Neuroscience. 21 (5), 696-706 (2018).
- Gibson, E. M., et al. Neuronal activity promotes oligodendrogenesis and adaptive myelination in the mammalian brain. Science. 344 (6183), 1252304 (2014).
- Pan, S., Mayoral, S. R., Choi, H. S., Chan, J. R., Kheirbek, M. A. Preservation of a remote fear memory requires new myelin formation. Nature Neuroscience. 23 (4), 487-499 (2020).
- Thornton, M. A., Hughes, E. G. Neuron-oligodendroglia interactions: Activity-dependent regulation of cellular signaling. Neuroscience Letters. 727, 134916 (2020).
- Ettle, B., Schlachetzki, J. C. M., Winkler, J. Oligodendroglia and myelin in neurodegenerative diseases: more than just bystanders. Molecular Neurobiology. 53 (5), 3046-3062 (2016).
- Essayan-Perez, S., Zhou, B., Nabet, A. M., Wernig, M., Huang, Y. A. Modeling Alzheimer's disease with human iPS cells: advancements, lessons, and applications. Neurobiology of Disease. 130, 104503 (2019).
- Li, L., et al. GFAP mutations in astrocytes impair oligodendrocyte progenitor proliferation and myelination in an hiPSC model of Alexander disease. Cell Stem Cell. 23 (2), 239-251 (2018).
- Lin, Y. T., et al. APOE4 causes widespread molecular and cellular alterations associated with Alzheimer's disease phenotypes in human iPSC-derived brain cell types. Neuron. 98 (6), 1294 (2018).
- TCW, J., et al. Cholesterol and matrisome pathways dysregulated in human APOE ε4 glia. bioRxiv. , (2019).
- Ang, C. E., Wernig, M.
- Penney, J., Ralvenius, W. T., Tsai, L. H. Modeling Alzheimer's disease with iPSC-derived brain cells. Molecular Psychiatry. 25 (1), 148-167 (2020).
- Zhang, Y., et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron. 78 (5), 785-798 (2013).
- Huang, Y. A., Zhou, B., Nabet, A. M., Wernig, M., Sudhof, T. C. Differential signaling mediated by ApoE2, ApoE3, and ApoE4 in human neurons parallels Alzheimer's Disease risk. Journal of Neuroscience. 39 (37), 7408-7427 (2019).
- Huang, Y. A., Zhou, B., Wernig, M., Sudhof, T. C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Abeta Secretion. Cell. 168 (3), 427-441 (2017).
- Yang, N., et al. Generation of oligodendroglial cells by direct lineage conversion. Nature Biotechnology. 31 (5), 434-439 (2013).
- Douvaras, P., et al. Efficient generation of myelinating oligodendrocytes from primary progressive multiple sclerosis patients by induced pluripotent stem cells. Stem Cell Reports. 3 (2), 250-259 (2014).
- Lee, E. H., Park, C. H. Comparison of reprogramming methods for generation of induced-oligodendrocyte precursor cells. Biomolecules & Therapeutics (Seoul). 25 (4), 362-366 (2017).
- Ehrlich, M., et al. Rapid and efficient generation of oligodendrocytes from human induced pluripotent stem cells using transcription factors. Proceedings of the National Academy of Sciences of the United States of America. 114 (11), 2243-2252 (2017).
- Rodrigues, G. M. C., et al. Defined and scalable differentiation of human oligodendrocyte precursors from pluripotent stem cells in a 3D culture system. Stem Cell Reports. 8 (6), 1770-1783 (2017).
- Hu, B. Y., Du, Z. W., Li, X. J., Ayala, M., Zhang, S. C. Human oligodendrocytes from embryonic stem cells: conserved SHH signaling networks and divergent FGF effects. Development. 136 (9), 1443-1452 (2009).
- Izrael, M., et al. Human oligodendrocytes derived from embryonic stem cells: Effect of noggin on phenotypic differentiation in vitro and on myelination in vivo. Molecular and Cellular Neuroscience. 34 (3), 310-323 (2007).
- Yamashita, T., et al. Differentiation of oligodendrocyte progenitor cells from dissociated monolayer and feeder-free cultured pluripotent stem cells. PLoS One. 12 (2), 0171947 (2017).
- Wang, S., et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell. 12 (2), 252-264 (2013).
- Chanoumidou, K., Mozafari, S., Baron-Van Evercooren, A., Kuhlmann, T. Stem cell derived oligodendrocytes to study myelin diseases. Glia. 68 (4), 705-720 (2020).
- Chetty, S., et al. A simple tool to improve pluripotent stem cell differentiation. Nature Methods. 10 (6), 553-556 (2013).
- Li, J., et al. A transient DMSO treatment increases the differentiation potential of human pluripotent stem cells through the Rb family. PLoS One. 13 (12), 0208110 (2018).
- Sambo, D., Li, J., Brickler, T., Chetty, S. Transient treatment of human pluripotent stem cells with DMSO to promote differentiation. Journal of Visualized Experiments: JoVE. (149), (2019).
- Douvaras, P., Fossati, V. Generation and isolation of oligodendrocyte progenitor cells from human pluripotent stem cells. Nature Protocols. 10 (8), 1143-1154 (2015).
- Mei, F., et al. Micropillar arrays as a high-throughput screening platform for therapeutics in multiple sclerosis. Nature Medicine. 20 (8), 954-960 (2014).
- Madhavan, M., et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nature Methods. 15 (9), 700-706 (2018).
- Zhang, Y., et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron. 89 (1), 37-53 (2016).
- Grubman, A., et al. A single-cell atlas of entorhinal cortex from individuals with Alzheimer's disease reveals cell-type-specific gene expression regulation. Nature Neuroscience. 22 (12), 2087-2097 (2019).
- Goldman, S. A., Kuypers, N. J. How to make an oligodendrocyte. Development. 142 (23), 3983-3995 (2015).
- Behrendt, G., et al. Dynamic changes in myelin aberrations and oligodendrocyte generation in chronic amyloidosis in mice and men. Glia. 61 (2), 273-286 (2013).
- Patzke, C., et al. Neuromodulator signaling bidirectionally controls vesicle numbers in human synapses. Cell. 179 (2), 498-513 (2019).
- Piao, J., et al. Human embryonic stem cell-derived oligodendrocyte progenitors remyelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell. 16 (2), 198-210 (2015).
- Keirstead, H. S., et al. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants remyelinate and restore locomotion after spinal cord injury. Journal of Neuroscience. 25 (19), 4694-4705 (2005).
- Kim, D. S., et al. Rapid generation of OPC-like cells from human pluripotent stem cells for treating spinal cord injury. Experimental & Molecular Medicine. 49 (7), 361 (2017).
Tags
Neurowetenschappen geïnduceerde pluripotente stamcellen iPS-cellen menselijke embryonale stamcellen ES-cellen neuronen oligodendrocyten oligodendrocyten voorlopercellen OPC's de ziekte van Alzheimer amyloïde-bètapeptiden synapsenErratum
Formal Correction: Erratum: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions
Posted by JoVE Editors on 12/29/2020.
Citeable Link.
An erratum was issued for: Generation of Human Neurons and Oligodendrocytes from Pluripotent Stem Cells for Modeling Neuron-Oligodendrocyte Interactions. The Representative Results section has been updated.
Figure 3 was updated from:
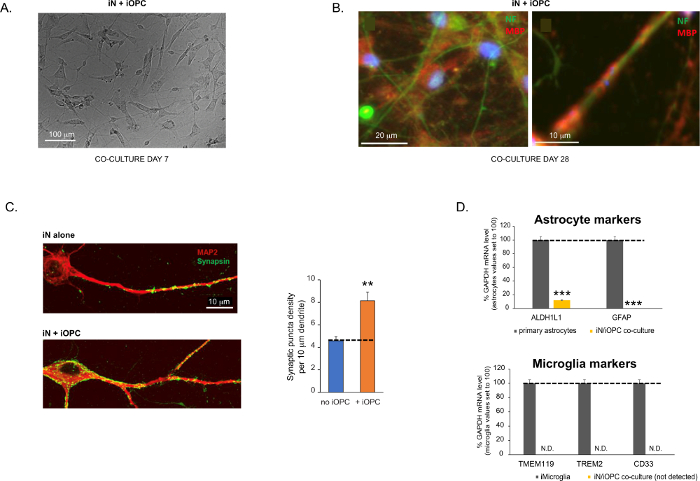
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. Please click here to view a larger version of this figure.
to:
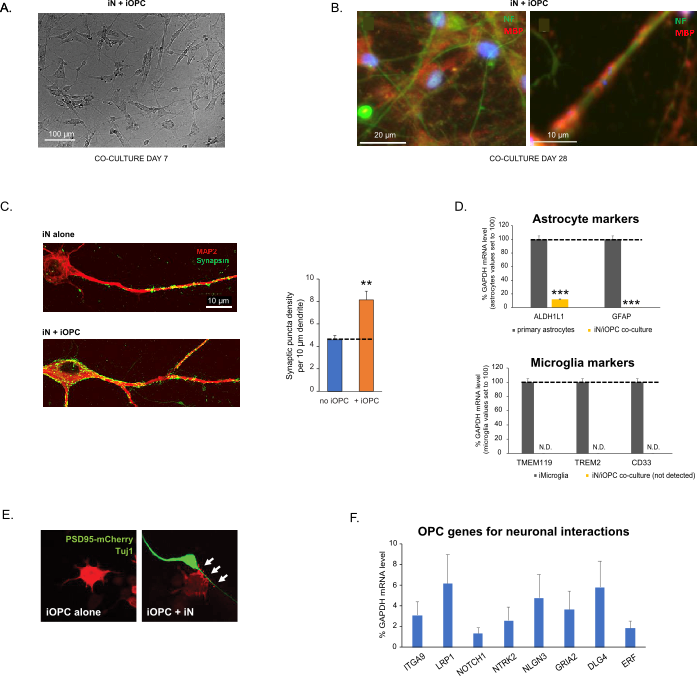
Figure 3: Co-culture of iNs and iOPCs. (A) Representative bright field image of co-cultured iNs and iOPCs at Day 7, showing a proper density for further maturation. (B) Representative immunofluorescence image of iNs and iOPCs co-cultured for 28 days. Axonal marker neurofilament NF is shown in green and oligodendrocytic marker MBP in red. Right, a segment of iN axon ensheathed by iOL process (MBP+). (C) Synapse formation assayed in 4-week-old co-cultures. Cells were stained for Synapsin 1 (Syn1, green) and MAP2 (red), and synaptic puncta were quantified by confocal analysis of density along the dendritic segments as described17,18. (D) In our co-cultures of iNs and iOPCs (7 days of co-culturing), the expression of astrocyte markers, ALDHL1 and GFAP, is minimal (top), and the expression of microglia markers, TMEM119, TREM2, and CD33, is not detected (N.D.) by qPCR. The contamination from these two glial cell types is thus excluded. (E) Coculturing iOPC with iN leads to the formation of neuron-OPC synapses. The fluorescence-tagged post-synaptic marker PSD95-mCherry is expressed only in OPCs, and display a diffuse pattern in single cultures (left) but aggregate to form puncta in cocultures (right, indicated by arrows; Tuj1, neuronal marker). (F) The expression of well-characterized oligodendroglial genes that can sense and respond to neuronal activities in the pure cultures of iOPCs at Day 14. Please click here to view a larger version of this figure.
The fourth paragraph was updated from:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D).
to:
Co-culturing of iNs and iOPCs
This protocol is optimized specifically for co-culturing iNs and iOPCs and allow our real-time monitoring of the inter-cellular communications between these two cell types along the course of neural development. The ideal plating densities for both cell types need to be decided with a series of cell number titration to achieve proper differentiation (Figure 3A). After 4 weeks in co-cultures, the iOPCs are expected to be adequately differentiated into OLs that are positive for specific markers such as MBP and extend processes to ensheath axons (Figure 3B). The co-culture system can robustly boost up the number of synapses, indicating that the iOPCs provide a neuronal support through physical contacts or release of trophic factors (Figure 3C). We can maintain the co-cultures in acceptable health condition for up to 6 weeks and observe that the synapse number and other neuronal attributes plateau around the fifth week. Of note, astrocytes and microglia are not present in our preparations and their absence can be documented by checking the expression of specific markers (Figure 3D). The iOPCs express a good number of well-characterized genes that can potentially respond to and mediate the activity-dependent signals from neighboring neurons, in a paracrine (e.g. neurotrophins and metabolites) and/or a synaptic manner (Figure 3E and 3F).
