Examples of electrographic seizure-like discharge recorded in the forebrain of an agar-embedded zebrafish larvae are shown in Figure 1. Large-amplitude multi-spike burst discharge in these samples was evoked by bath application of a convulsant drug, 40 mM pilocarpine (in A; 6 dpf) or 1 mM picrotoxin (in B; 8 dpf). In these recordings, immobilized and agar-embedded zebrafish are continuously monitored for up to 90 min. Fish remain viable under these recording conditions for up to 24 hr. Drugs are added to the bathing medium and normally diffuse into the agar to elicit activity in the larval zebrafish forebrain within 30 to 45 min. This method can be used with relatively small volumes of drug solution (2-5 ml) as larvae do not require continuous perfusion and can be recorded in a static bath configuration if necessary. In panel C, a spontaneous burst discharge is shown for a genetically modified zebrafish at 3 dpf; recording was made in zebrafish recording media. Morpholino oligonucleotide injection at the 1-2 cell stage was used to knockdown expression of a gene for Tuberous Sclerosis Complex (tsc1a), a pediatric form of epilepsy associated with seizures and autism. Extracellular recordings can also be obtained from the optic tectum. For examples, see Baraban et al. (2005)9 or Baraban et al. (2007)10. As shown here, the electrical events can vary in waveform and duration depending on the mechanism of action used to elicit seizure activity.
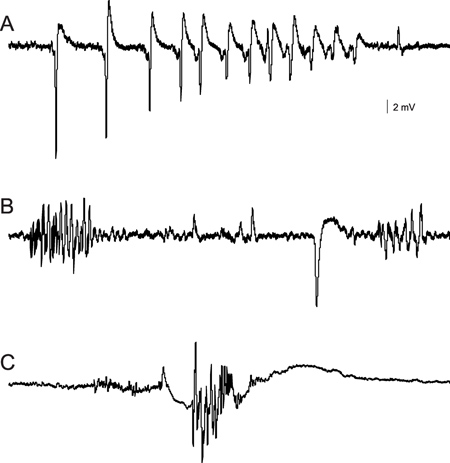
Figure 1. Extracellular field recordings of abnormal burst discharge activity recorded in the forebrain of immobilized and agar-embedded zebrafish larvae. Recording electrodes are positioned under visual observation on an Olympus BX50 upright microscope. Recordings in (A) and (B) were initiated approximately 40 to 45 min after drug application. (A) Multi-spike discharge recorded in pilocarpine, a muscarinic acetylcholine receptor antagonist. (B) Burst discharges recorded in picrotoxin, a GABA-A receptor antagonist. (C) A single multi-spike burst discharge recorded from a 3 dpf zebrafish larvae injected with a morpholino against tsc1a.
