Isolation and immunopurification of villous cytotrophoblast cells from a normal term placenta obtained by vaginal delivery yielded 1 x 108 viable cells. The placenta weighed 350 g, was 19 cm in diameter, 4 cm tall with discoid shape and transparent membranes. No cotyledon malformation was detected. The umbilical cord had paracentral localization and a length of 56 cm. The purity was evaluated by flow cytometry using vimentin and cytokeratin-7 markers. More than 98% of the cells were negative for vimentin and positive for cytokeratin-7, confirming the purity of villous trophoblasts cells obtained from the immunopurification. Villous cytotrophoblast cells were added to 96-well culture plates under normoxic conditions in the presence or absence of 1 mM melatonin. The biochemical differentiation of villous cytotrophoblasts was monitored by determining levels of β-human chorionic gonadotropin (β-hCG) secretion as described previously1,7,20,21. The morphological differentiation and apoptosis were assessed by immunofluorescence using anti-desmoplakin and anti-caspase-cleaved cytokeratin 18 intermediate filaments7,22. Cell culture media from day 1 (mainly villous cytotrophoblasts) to day 4 (mainly syncytiotrophoblasts) were collected, centrifuged and β-hCG levels were measured in the supernatants. Production of β-hCG, which is exclusive to the syncytiotrophoblast, increased with culture time (Figure 4). Not only hypoxia/reoxygenation, but hyperoxia (> 20% O2) also activated apoptosis23. Thus, adoption of an 8% O2 concentration was representative of the quantity of oxygen to which a villous trophoblast cell would be exposed during the third trimester of pregnancy10. The peak of β-hCG levels observed at 72 hr confirmed the capacity of villous cytotrophoblasts to differentiate under these conditions. Melatonin did not alter β-hCG secretion under these study conditions. The decrease of β-hCG levels at 96 hr was likely caused by apoptosis of trophoblast cells, which increases after prolonged periods in primary culture5,7,22,24,25 (Figure 4). DMSO (0.1% vol/vol) was selected because it did not affect β-hCG levels26,27. The protective role of melatonin was strongly related to its antioxidant properties. Hypoxia/reoxygenation after 72h of culture induced oxidative stress in villous trophoblast cells. The protective effect of melatonin was assessed with Reactive Oxygen Species (ROS) Detection Reagent (Figure 5A). After 96 hr of culture, trophoblast cells were incubated for 45 min with 10 µM of 5-(and-6)-carboxy-2′,7′-dichlorodihydrofluorescein diacetate (carboxy-H2DCFDA) to detect the total amount of ROS produced8. Villous trophoblast cells that underwent hypoxia/reoxygenation had significantly increased ROS levels (54%) compared to those under normoxia. This increase was reversed by treatment with 1 mM melatonin. Moreover, under normoxia melatonin did not modulate ROS levels (homeostasis), which was similar to non-treated villous trophoblast cells (Figure 5A). Figure 4 and 5A show that under normoxia melatonin did not alter levels of oxidative stress or β-hCG secretion in the trophoblast cells, which corroborates previous studies showing no modulation of cell homeostasis under normal conditions28,29.
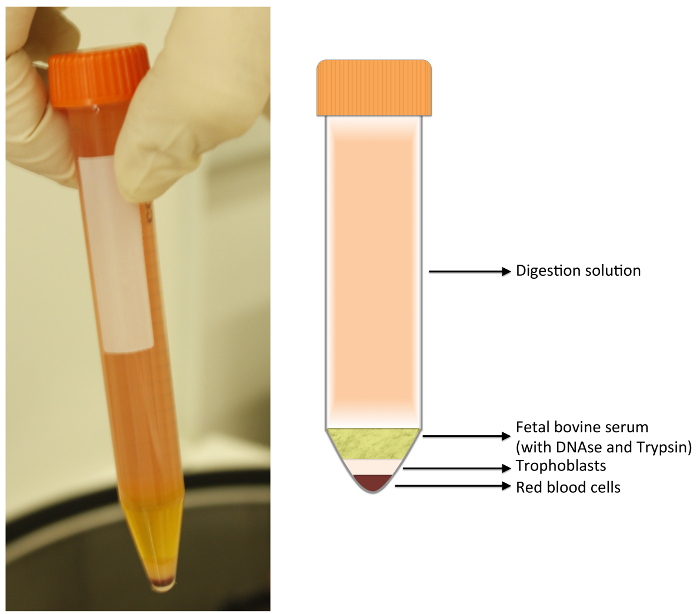
Figure 1: Digestion Tube. After centrifugation, 4 layers are formed. The upper layer is composed of digestion solution; just below, the fetal bovine serum (FBS). Both layers should be discarded with a vacuum pump. The lower layers are composed as follow: a white layer containing fibroblasts, leukocytes, macrophages, and trophoblasts; and a bottom layer composed of red blood cells. Please click here to view a larger version of this figure.
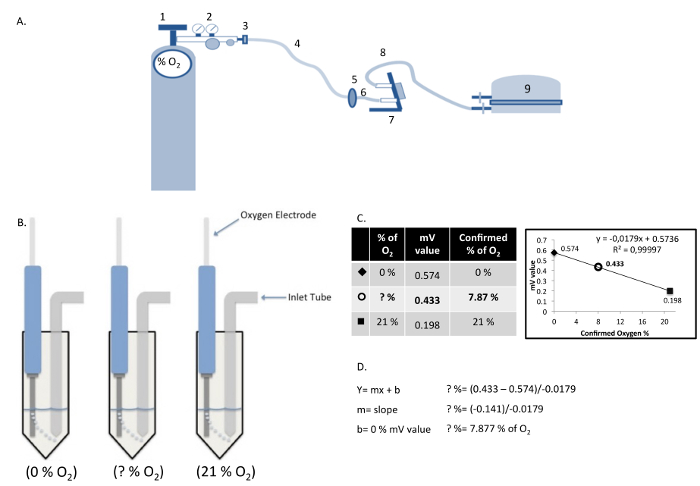
Figure 2: Components of the Hypoxia Chamber and Measurement/Calculation of Dissolved Oxygen Concentration. (A) Hypoxia chamber and gas cylinder assembly: (1) Gas cylinder; (2) Gas regulator; (3) Gas hose clamp; (4) Cylinder gas hose; (5) Inlet filter; (6) Inlet hose; (7) Flow meter; (8) Outlet hose; (9) Modular incubator chamber. (B) Calculation of actual oxygen concentration in cell culture medium using a standard curve produced with known oxygen concentrations. (C and D) The relative values obtained in the solutions "0% O2" and "21% O2", are plotted graphically as a linear function to determine the oxygen concentration in the cell culture medium "?% O2". Please click here to view a larger version of this figure.
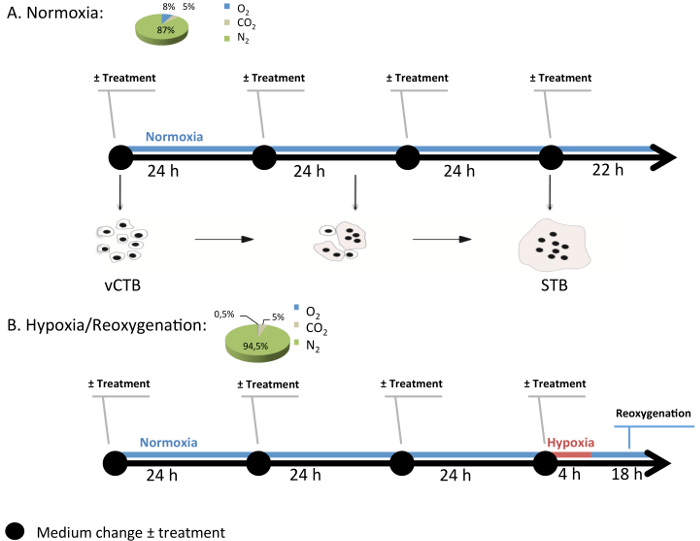
Figure 3: Generic Experimental Design of Cell Culture in the Modular Incubation Chamber. Normoxia (8% O2; 5% CO2; 87% N2) and hypoxia/reoxygenation (H/R) (0.5% O2; 5% CO2; 94.5% N2) are conditions used to study pathological conditions in villous cytotrophoblast (vCTB) and syncytiotrophoblast (STB) cells. Every 24 hr, medium with or without melatonin (1 mM) is changed and the gas mixture is renewed. Under H/R, STB cells undergo hypoxia (0.5% O2) for 4 hr and then return to normoxia (8% O2). Please click here to view a larger version of this figure.
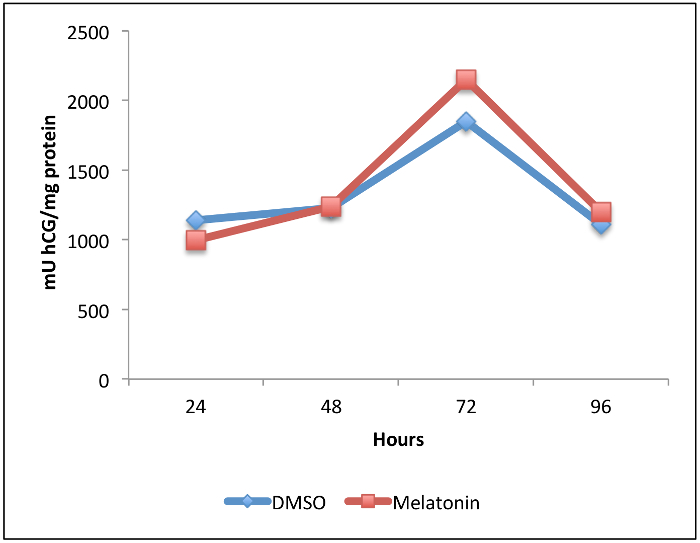
Figure 4: Effect of Melatonin on beta-human Chorionic Gonadotropin (β-hCG) Secretion during Villous Trophoblast Differentiation. Villous cytotrophoblast cells were isolated and purified from human healthy term placentas. Cells were treated for 96 hr with 1 mM melatonin or dimethyl sulfoxide (DMSO 0.1%: vehicle control) under normoxic conditions (8% O2; 5% CO2; 87% N2). β-hCG levels in culture medium were measured by enzyme-linked immunosorbent assay (ELISA) after 24, 48, 72 and 96 hr of primary culture. Levels were normalized to the protein content of the whole-cell lysate from each corresponding well. Please click here to view a larger version of this figure.
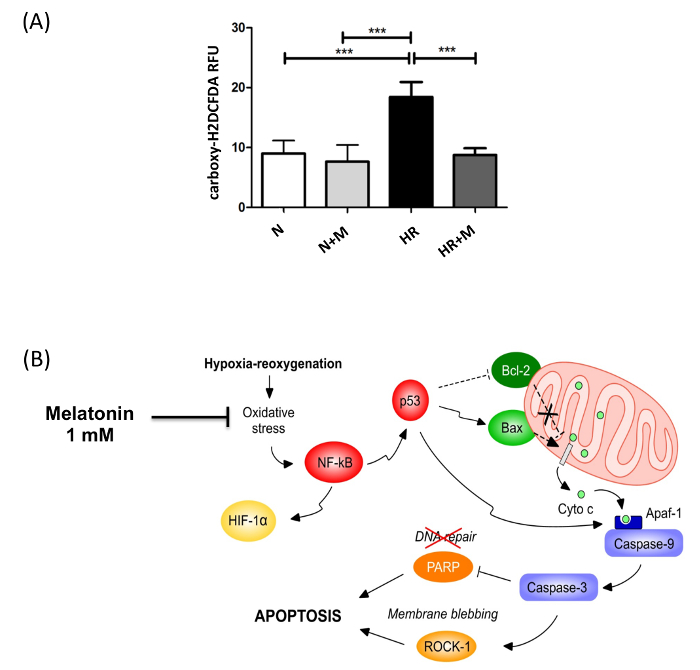
Figure 5: Anti-oxidant Effect of Melatonin in Syncytiotrophoblast Exposed to Hypoxia/Reoxygenation. (A) The effect of melatonin (M; 1 mM) on intracellular reactive oxygen species (ROS) levels in syncytiotrophoblast cells under normoxia (N) or hypoxia/reoxygenation (HR), induced after 72 hr of culture, was assessed by 5-(and-6)-carboxy-2′,7′-dichlorodihydrofluorescein diacetate (carboxy-H2DCFDA) fluorescence. Results are expressed as the mean ± SD of 3 different placentas; *** P < 0.001 (Lanoix, et al.8). (B) The cellular pathways involved in the putative protection of melatonin against hypoxia/reoxygenation-induced apoptosis. Primary villous cytotrophoblast cells were cultured for 72 hr under normoxia (8% O2) to allow differentiation into syncytiotrophoblast. Cells were exposed to 1 mM of melatonin or vehicle control and then subjected to hypoxia (0.5% O2) for 4 hr followed by an 18 hr reoxygenation period (8% O2). Hypoxia/reoxygenation-induced oxidative stress activates redox sensitive transcription factors such as nuclear factor kappa B (NF-κB) and hypoxia inducible factor 1 (HIF-1). NF-κB induces p53, which triggers the Bax/Bcl-2 pathway of mitochondrial apoptosis involving the cleavage and activation of caspases 9 and 3. Caspase 3 activates Rho-associated, coiled coil-containing protein kinase 1 (ROCK-1), the cleavage of poly(ADP-ribose) polymerase (PARP) and the impairment of DNA repair. Melatonin prevents the induction of mitochondrial apoptosis by acting as a powerful antioxidant to reduce the oxidative stress caused by hypoxia/reoxygenation. This figure has been modified from Lanoix et al., 20138. Please click here to view a larger version of this figure.
| Digestion 1 | Digestion 2 | Digestion 3 | Digestion 4 | |
| Modified HBSS (ml) | 150 | 100 | 75 | 75 |
| DNAse (µl) | 300 | 200 | 150 | 150 |
| (0.1 mg/µl) | ||||
| MgSO4 (µl) | 150 | 100 | 75 | 75 |
| (800 mM) | ||||
| CaCl2 (µl) | 150 | 100 | 75 | 75 |
| (100 mM) | ||||
| Trypsin (U) | 1,824,000 | 1,200,000 | 960,000 | 960,000 |
| P/S (ml) | 1 | 0 | 0 | 0 |
Table 1: Quantities of Ingredients for the Digestion Solution. Penicillin and streptomycin (P/S); magnesium sulfate (MgSO4); calcium chloride (CaCl2); deoxyribonuclease IV (DNase IV).
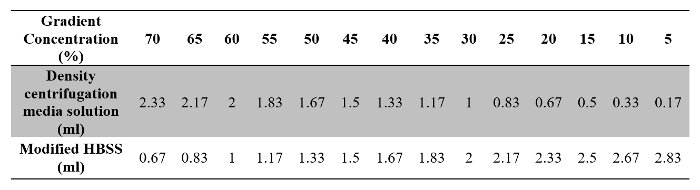
Table 2: Volumes of Density Centrifugation Media Solution and Modified HBSS Required for Preparation of the Gradient Solution.
