Peripheral injection of alpha-synuclein prionoids via the tongue or the peritoneum induced neuropathology in the CNS of bigenic Tg(M83+/-:Gfap-luc+/-) mice (Table 1 and Figure 1). After a single intraperitoneal injection with alpha-synuclein fibrils, four of five mice developed neurologic disease with a median incubation time of 229 ±17 days. Surprisingly, only one of five mice developed CNS disease after intraglossal injection with alpha-synuclein fibrils after 285 days. Inoculation with PBS did not cause disease within the 420 day period of the experiment.
Biochemical analysis by Western blotting and probing with the phospho-specific EP1536Y antibody against Ser129 of alpha-synuclein revealed that for both transmission routes diseased animals had accumulated aggregates of phosphorylated and sarkosyl-insoluble alpha-synuclein in their brains and spinal cords (Table 2 and Figure 2). In contrast, brains and spinal cords of PBS-injected and non-diseased animals only contained monomeric species of phosphorylated alpha-synuclein.
Immunofluorescent staining of brain slices of intraperitoneally injected and diseased mice with the phospho-specific pSyn#64 antibody revealed a wide distribution of phosphorylated alpha-synuclein deposits throughout the CNS (Table 2 and Figure 3). Moreover, phosphorylated alpha-synuclein deposits colocalized with ubiquitin and p62 indicating an aberrant protein homeostasis that might contribute to the formation of deposits.
The accumulation of alpha-synuclein aggregates in diseased animals was accompanied by neuroinflammatory changes, which was detected by immunofluorescence staining for GFAP, a marker of astrocytes that can reveal reactive astrogliosis (Figure 4). Additionally, activated microglia were also found in regions with abundant alpha-synuclein pathology by immunofluorescence staining for IBA-1 (ionized calcium-binding adapter molecule 1). In accordance, bioluminescence imaging revealed increased radiance (>2 × 106 p/s/cm2/sr) emitted from the brains of Tg(M83+/-:Gfap-luc+/-) mice injected with alpha-synuclein fibrils, shortly before they developed signs of neurologic disease. In contrast, brains of mice injected with PBS did not show any increase in radiance. Increased radiance is indicative of reactive astrogliosis, a hallmark of neuroinflammation, since luciferase expression is driven from the Gfap promoter.
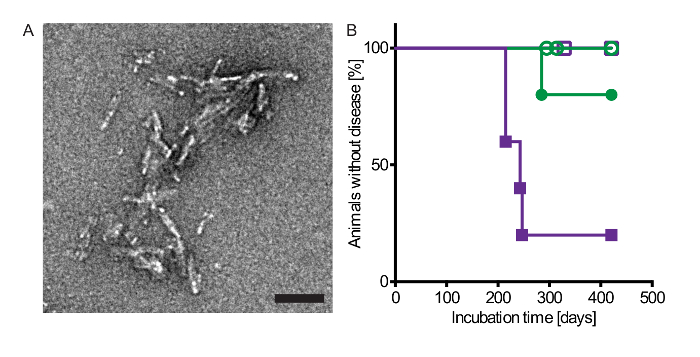
Figure 1: Injection with alpha-synuclein fibrils via the intraperitoneal or intraglossal route causes disease in bigenic Tg(M83+/-:Gfap-luc+/-) mice. (A) Electron micrograph of sonicated recombinant human alpha-synuclein fibrils that were intraperitoneally and intraglossally injected into mice. Negative staining with uranyl acetate revealed multiple clusters of short, rod-shaped aggregates. Scale bars = 100 nm. (B) Kaplan-Meier survival curves show that after intraperitoneal injection with alpha-synuclein fibrils four of five Tg(M83+/-:Gfap-luc+/-) mice developed neurologic disease in 229 ±17 days (purple closed squares), whereas none of the PBS-injected control mice developed signs of neurologic dysfunction within 420 days (purple open squares). After intraglossal inoculation with alpha-synuclein fibrils one mouse of five died after 285 days (green closed circles). In contrast, none of the PBS-injected control mice developed neurologic disease or spontaneously died (green open circles). Modified from Breid et al., 201614. Please click here to view a larger version of this figure.
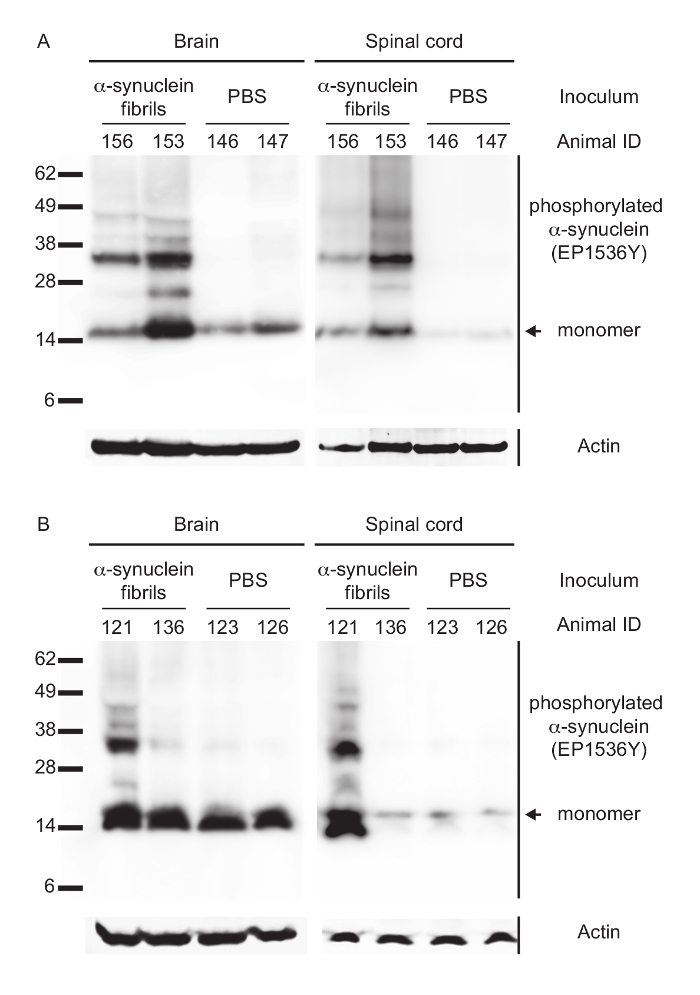
Figure 2: Biochemical analysis of phosphorylated alpha-synuclein in the CNS of peripherally injected Tg(M83+/-:Gfap-luc+/-) mice. (A) Immunoblotting with the EP1536Y antibody, recognizing phosphorylation at Ser129 of alpha-synuclein, showed that intraperitoneally injected and diseased mice accumulated high-molecular-weight species of sarkosyl-insoluble aggregates of phosphorylated alpha-synuclein in their brains and spinal cords. Control mice challenged with PBS did not accumulate aggregates of phosphorylated alpha-synuclein and showed bands only for its monomeric form. (B) After intraglossal challenge with alpha-synuclein fibrils only one mouse, animal 121, showed sarkosyl-insoluble aggregates of phosphorylated alpha-synuclein in its brain and spinal cord, whereas all other intraglossally inoculated mice remained healthy without deposits of alpha-synuclein. Molecular masses are shown in kilodaltons. Sample loading in each lane is shown by detection of actin. Modified from Breid et al., 201614. Please click here to view a larger version of this figure.
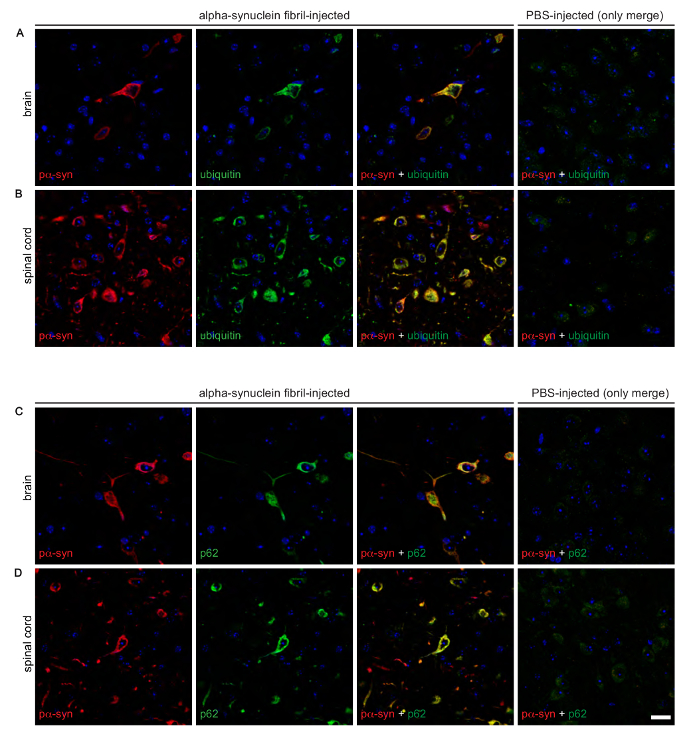
Figure 3: Immunofluorescence analysis shows that deposits of phosphorylated alpha-synuclein colocalize with ubiquitin and p62 in the brains and spinal cords of diseased Tg(M83+/-:Gfap-luc+/-) mice. Co-staining of phosphorylated alpha-synuclein, detected with the EP1536Y antibody, and ubiquitin revealed a distinct colocalization in the brains, as observed in the thalamic nucleus (A), and spinal cords, as detected in the grey matter (B), of diseased mice after intraperitoneal challenge with alpha-synuclein fibrils. Similarly, phosphorylated alpha-synuclein, detected with the pSyn#64 antibody, also colocalized with p62 in brains (C) and spinal cords (D) of diseased animals. (A to D) PBS-injected healthy control animals did not show deposits or colocalization for any of these proteins. Nuclear staining with DAPI is shown in blue. Scale bars = 20 µm. Modified from Breid et al., 201614. Please click here to view a larger version of this figure.
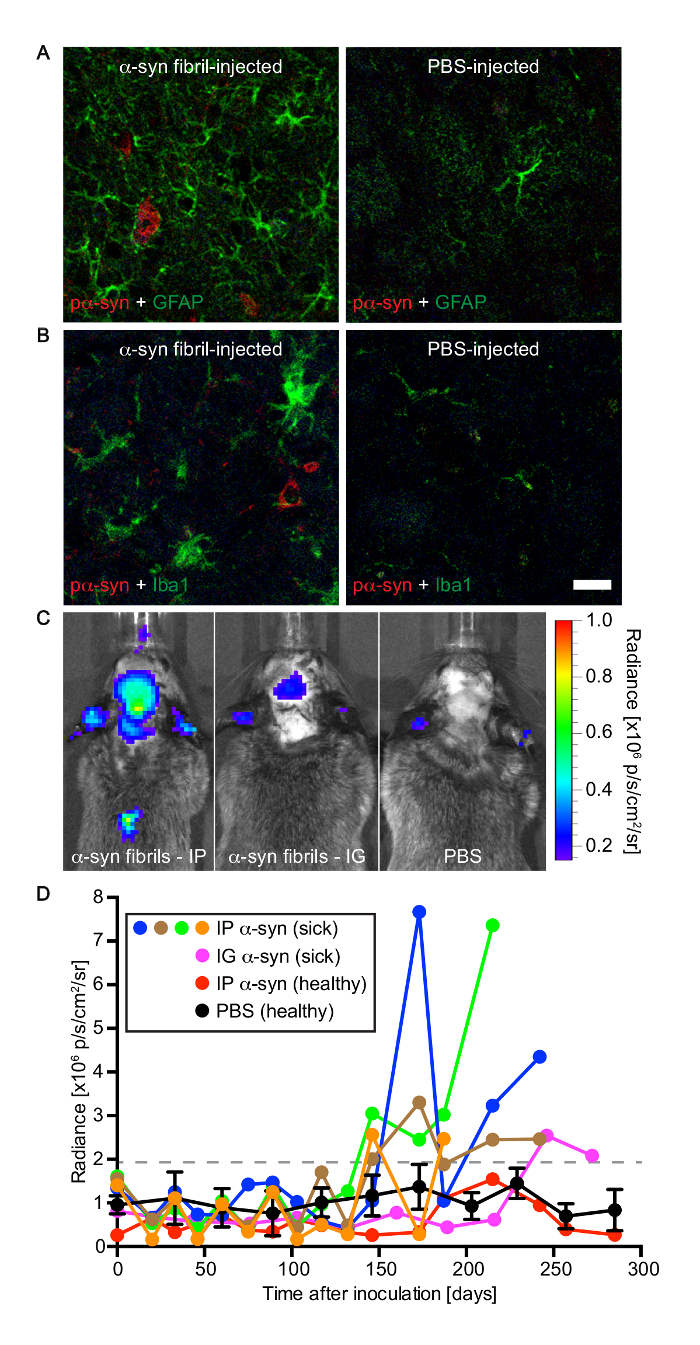
Figure 4: Tg(M83+/-:Gfap-luc+/-) mice developed gliosis after peripheral challenge with alpha-synuclein fibrils. (A) Immunofluorescence analysis of brain sections of diseased animals with an antibody against GFAP, a marker of astrocytes, revealed reactive astrogliosis in areas with deposits of phosphorylated alpha-synuclein, which were detected in the brain stem with the pSyn#64 antibody. In contrast, there was no reactive astrogliosis in brains of PBS-injected control mice. (B) Similarly, staining with an antibody to IBA-1, a marker of microglia, demonstrated microgliosis in areas with deposits of phosphorylated alpha-synuclein in diseased animals. Brains of healthy, PBS-injected control mice did not show signs of microgliosis. (C) Bioluminescence imaging revealed elevated radiance from the brains and spinal cords of Tg(M83+/-:Gfap-luc+/-) mice intraperitoneally injected with alpha-synuclein fibrils (left panel), which was caused by the activation of astrocytes, shortly before the mice developed neurologic symptoms. In the brains and spinal cords of PBS-injected control mice the basal radiance did not increase with time (right panel). After intraglossal inoculation with alpha-synuclein fibrils, one Tg(M83+/-:Gfap-luc+/-) mouse, animal 121, showed signs of reactive astrogliosis shortly before it died at 285 days (center panel). (D) After intraperitoneal challenge of Tg(M83+/-:Gfap-luc+/-) mice with alpha-synuclein fibrils, increased levels of bioluminescence (>2 × 106 p/s/cm2/sr) were measured from the brains of four mice (blue, green, brown, and orange circles) some weeks before they developed neurologic signs of disease. One of five animals also showed elevated levels of bioluminescence shortly before it died 285 days after intraglossal injection with alpha-synuclein fibrils (magenta circles). In contrast, for healthy, PBS-injected control mice (black circles; error bars show SD [n = 4]) and one animal that did not develop disease after intraperitoneal injection with alpha-synuclein fibrils (red circles), the bioluminescence signal remained below a threshold of 2 x 106 p/s/cm2/sr within the period of observation. Scale bars = 20 µm. Modified from Breid et al., 201614. Please click here to view a larger version of this figure.
| Mouse line | Inoculum (μg) | Inoculation route | No. of mice with disease/no. of mice inoculated | Mean survival time ±SD (days) |
| Tg(M83+/-:Gfap-luc+/-) | human α-synuclein fibrils (50) | intraperitoneal | 4/5 | 229 ±17/420 |
| Tg(M83+/-:Gfap-luc+/-) | PBS | intraperitoneal | 0/5 | 0/420 |
| Tg(M83+/-:Gfap-luc+/-) | human α-synuclein fibrils (10) | intraglossal | 1/5 | 285/420 |
| Tg(M83+/-:Gfap-luc+/-) | PBS | intraglossal | 0/4 | 0/420 |
Table 1: Inoculation experiments. *Modified from Breid et al., 201614
| Target [antibody clone] | Host | Immunogen | Dilution in IF | Dilution in WB |
| Actin [C4] | Mouse | – | – | 1:1,000 |
| α-synuclein, phospho S129 [pSyn#64] | Mouse | pSer129 | 1:1200 | – |
| α-synuclein, phospho S129 [EP1536Y] | Rabbit | pSer129 | 1:100 | 1:1,000 |
| Glial fibrillary acidic protein (GFAP) | Rabbit | – | 1:200 | – |
| IBA-1 | Rabbit | – | 1:500 | – |
| Sequestosome-1 (p62) | Rabbit | – | 1:100 | – |
| Ubiquitin [Ubi-1] | Mouse | – | 1:500 | – |
Table 2: Antibodies used for immunofluorescence (IF) and Western blotting (WB). *Modified from Breid et al., 201614
