In order to illustrate the expression levels that can be obtained by the intraspinal injection of rAAV encoding a marker protein, we first injected AAV1.CAG.eGFP into the lumbar spinal cord of wild-type mice. Three injections spaced approximately 1 mm apart produced a nearly continuous infection of lumbar spinal segments L3 to L5 (Figure 1A-C). Virus injection at a depth of 300 µm from the spinal surface leads to predominant infection of cells in the spinal cord dorsal horn. However, infected cells could also be found in the ventral horn (Figure 1D). Next, the goal was to illustrate the difference in rAAV-mediated expression when injecting a Cre-dependent rAAV into a Cre transgenic mouse. We therefore injected the Cre-dependent AAV1.CAG.flex.eGFP vector into the spinal cord of GlyT2::Cre transgenic mice. As before, eGFP expression was observed in the dorsal and ventral horn. However, as expected, the expression of eGFP became more restricted, reflecting the distribution of GlyT2+ neurons, i.e. the relatively sparse expression in the superficial dorsal horn and dense expression in the deep dorsal horn (Figure 1E).
Next, to demonstrate the feasibility of addressing circuit function of spinal neuronal subpopulations, two different Cre-dependent AAVs coding for different effector proteins (DTA and hM3Dq) were injected into GlyT2::Cre transgenic mice. Analogous to the viral expression observed after three injections of AAV1.CAG.eGFP into wild-type mice, there was robust ablation of inhibitory neurons (Pax2+) in lumbar segments L3-L5 after the injection of AAV1.EF1a.flex.DTA into Glyt2::Cre mice (Figure 2A,C). Loss of glycinergic inhibitory neurons in these segments evoked a marked mechanical hypersensitivity (Figure 2D) and spontaneous aversive behavior directed towards the ipsilateral hindlimb (Figure 2E). The aversive behavior led to self-inflicted lesions, which could be observed on the paws, calf, and thigh (for data, see Foster et al.7) Opposite effects were observed when activating glycinergic neurons through injection of AAV1.hSyn.flex.hM3Dq and subsequent intraperitoneal injection of CNO (Figure 2B). Mice became desensitized to noxious mechanical stimulation (Figure 2F) and other noxious stimuli (see Foster et al.7) In addition, when treated with the pruritogens histamine or chloroquine, hM3Dq-mediated activation of glycinergic neurons efficiently suppressed prurifensive responses (Figure 2G). These results demonstrate that three injections of rAAV into the lumbar spinal cord are able to transduce an area of the lumbar spinal cord sufficient to observe robust behavioral changes evoked by stimulation of the corresponding hindlimb.
In a final set of experiments, we wanted to demonstrate the potential differences in using Cre reporter mice compared to Cre reporter viruses. Therefore, a Cre driver gene was chosen that has previously been described as displaying a restricted expression pattern in the mouse spinal cord. The gene RORβ has been suggested to be expressed predominantly in inhibitory interneurons of the deep dorsal horn13,14. This study used RORβCre knock-in mice and crossed them to Rosa26lox-STOP-lox-tdTomato (R26Tom) Cre reporter mice, which lead to the expression of tdTomato in all cells displaying Cre expression at any time point before analysis (Figure 3A). Characterization of tdTomato+ cells in RORβCre; R26Tom mice revealed expression in neurons and astrocytes of the dorsal horn (Figure 3B,C). In fact, quantification of the tdTomato+ cells suggested that the majority of cells undergoing Cre-mediated recombination (58%) were non-neuronal. We then injected a Cre-dependent tdTomato reporter rAAV (AAV1.CAG.flex.tdTomato) into the spinal cord of P40 RORβCre mice (Figure 3D). In contrast to R26Tom-mediated reporter expression, we found all tdTomato+ cells labeled by the reporter virus to be neurons (Figure 3E,F). Finally, the identity of the tdTomato+ neurons was analyzed in both sets of mice (RORβCre; R26Tom and RORβCre injected with AAV1.CAG.flex.tdTomato). In both cases, the majority of neurons were inhibitory (>85%) and the minority of neurons excitatory (<20%) (Figure 3 G-I), which is in agreement with previous assessments of the identity of RORβ+ neurons13,14.
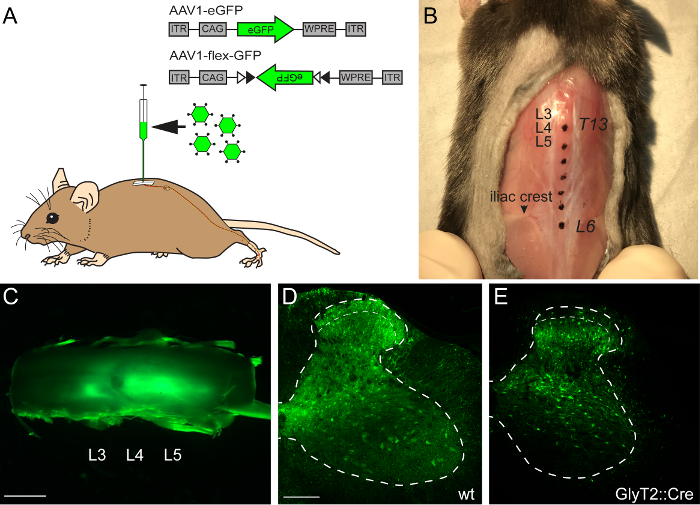
Figure 1: Intraspinal Injection of AAV1-eGFP/AAV1-flex-eGFP
(A) Schematic illustration of an intraspinal injection of an AAV1.CAG.eGFP (AAV1-eGFP) into the lumbar spinal cord, which is innervated from the hindlimb. (B) Anatomical location of the lumbar spinal cord segments L3-L4 can be seen in a top down view of the back of a mouse. The skin was opened to expose the vertebral column. Spinal processes of vertebrae T13-L6 are colored as anatomical references and an arrow indicates the iliac crest. (C) Representative image of a whole mount lumbar spinal cord. Green fluorescence indicates virus-transduced areas of the spinal cord. (D) Representative image of a cross section through an AAV1-eGFP-transduced lumbar spinal cord from a wild-type mouse. (E) Representative image of a cross section of an AAV1.CAG.flex.eGFP-transduced spinal cord of a GlyT2::Cre mouse. Dashed lines represent outline of the gray matter and superficial dorsal horn of the spinal cord. Scale bars C = 1 mm, D = 100 µm. Please click here to view a larger version of this figure.
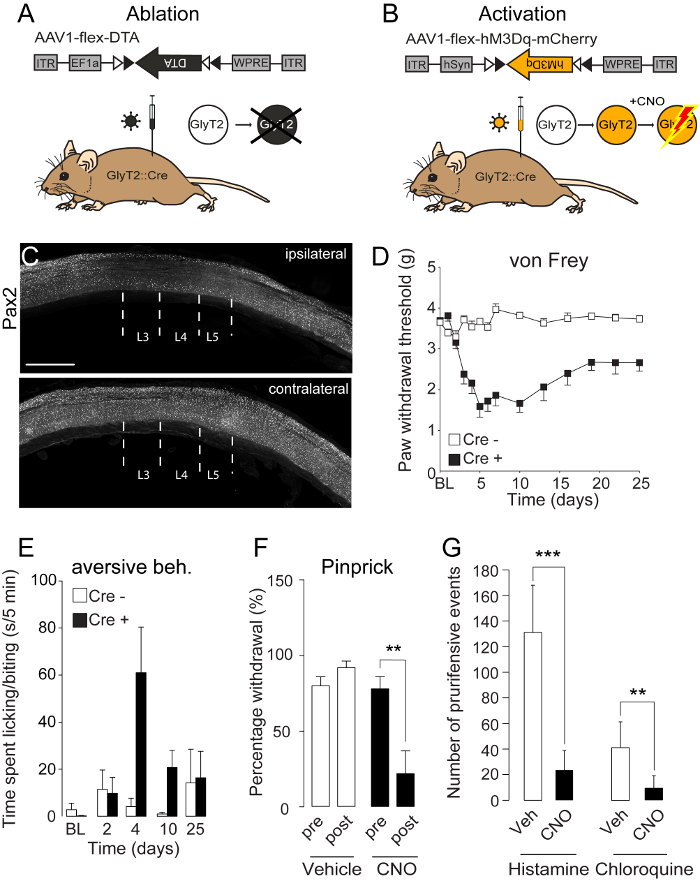
Figure 2: Functional Manipulation of Spinal Cre-expressing Glycinergic Neurons
(A+B) Schematic illustration of an intraspinal injection of an AAV1.EF1a.flex.DTA (A) or an AAV1.EF1a.flex.hM3Dq (B) into the lumbar spinal cord. Cre-dependent expression of diphtheria toxin fragment A (DTA) will lead to ablation of the glycinergic neurons (GlyT2+) (A), while Cre-dependent expression of the pharmacogenetic designer receptor hM3Dq will render GlyT2 neurons activatable by clozapine-N-oxide (CNO) (B). (C) Three injections of AAV1.EF1a.flex.DTA into the L3-L5 segments of GlyT2::Cre mice led to a marked loss of inhibitory neurons in the respective segments of the ipsilateral but not of the contralateral side. (D) Loss of GlyT2 neurons evoked a long-lasting hypersensitivity to mechanical von Frey stimulation in the ipsilateral hind paw of GlyT2::Cre mice injected with AAV1.EF1a.flex.DTA, but no change was observed if Cre-negative mice were injected. (E) Loss of GlyT2 neurons evoked spontaneous aversive behavior reminiscent of chronic itch. (F) hM3Dq-mediated activation of GlyT2 neurons alleviated noxious mechanical pain evoked by pinprick stimulation. (G) hM3Dq-mediated activation of GlyT2 neurons reduced pruritogen (chloroquine or histamine)-evoked aversive behavior. Data are represented as mean ± SEM. *** p < 0.001; ** p < 0.01. Scale bar C = 1 mm. g = grams. Images are re-used and modified from Foster et al.7 Please click here to view a larger version of this figure.
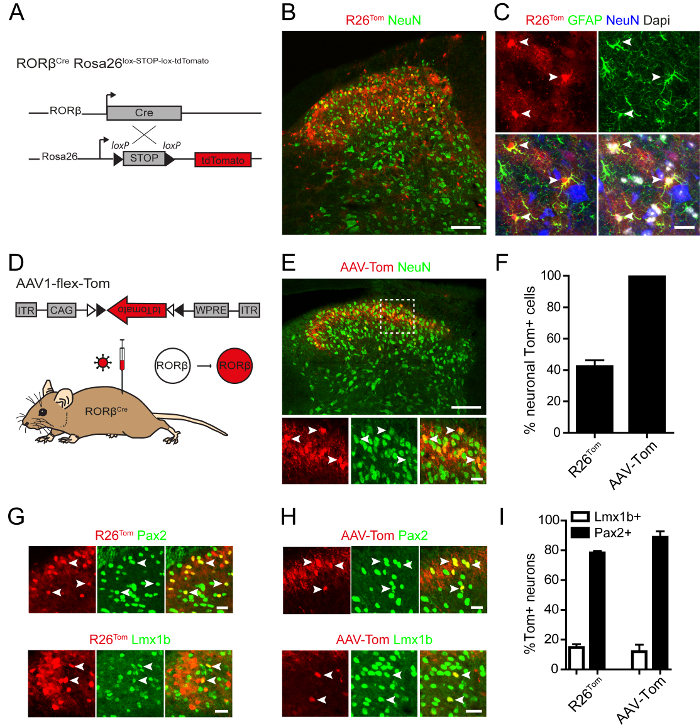
Figure 3: Genetic and Virus-mediated Labeling of RORβ-expressing Cells. (A) Diagram showing the strategy for Cre-dependent expression of fluorescent tdTomato reporter in RORβCre; Rosa26lox-STOP-lox-tdTomato (R26Tom) mice. (B) Immunostaining on spinal cord sections of RORβCre; R26Tom mice revealed that NeuN+ RORβ-Tom neurons can be found in laminae I-IV. (C) Cre-dependent expression of tdTomato was also observed in GFAP+ cells of RORβCre; R26Tom mice, suggesting that RORβ is expressed in astrocytes during development. Arrowheads indicate double-labeled astrocytes in lamina III. (D) Diagram showing intraspinal injection of AAV-flex-Tom (rAAV1.CAG.flex.tdTomato) virus into RORβCre mice to drive local Cre-dependent expression of tdTomato. (E) Immunostaining on spinal cord sections of RORβCre mice injected with AAV-flex-Tom virus. RORβ-Tom neurons were localized to the superficial laminae of the spinal dorsal horn and expression of tdTomato was absent from astrocytes. (F) Percentage of RORβ-Tom cells expressing the neuronal marker NeuN in spinal sections of RORβCre; R26Tom mice and RORβCre mice injected with AAV-flex-Tom virus. (G-H) Immunostaining on spinal cord sections of (G) RORβCre; R26Tom mice and (H) RORβCre mice injected with AAV-flex-Tom virus showing colocalization between RORβ-Tom neurons and the inhibitory marker Pax2 or the excitatory marker Lmx1b. (I) Percentage of RORβ-Tom neurons expressing Lmx1b and Pax2 in RORβCre; R26Tom mice and RORβCre mice injected with AAV-Tom virus. Data are represented as mean ± SEM. Data are from 2-3 mice and 1-3 sections per mouse. Scale bars represent 100 µm (B,E) and 20 µm (C,E in high magnification images, G and H). Please click here to view a larger version of this figure.
| Variable | Set value | Unit |
| heat (H) | 450 | – (value proportional to power of the radiated heat) |
| force preliminary pull (F(TH)) | 20 | – (value proportional to voltage applied to force coil) |
| distance threshold (s(TH)) | 25 | 0.12 mm |
| delay heatstop (t(H)) | 30 | 0.5 ms |
| distance heatstop (s(H)) | 0 | 0.12 mm |
| delay pull 1 (t(F1)) | 200 | 0.5 ms |
| force pull 1 (F1) | 300 | – (value proportional to voltage applied to force coil) |
| distance pull 2 (s(F2)) | 30 | 0.12 mm |
| force pull 2 (F2) | 600 | – (value proportional to voltage applied to force coil) |
| adjust (AD) | 0 | – |
Table 1: Puller Settings
