Specific neurons were genetically labeled with either EGFP or GCaMP6s, whose expression were driven in Gal4 lines. A Gal4 line gSAIzGFFM119B was used to label a nucleus in the pretectal area (magnocellular superficial pretectal nucleus), and a subpopulation of olfactory bulb neurons. Another Gal4 line, hspGFFDMC76A, was used to label the ILH. We laser-ablated the pretectal neurons bilaterally (Figure 2A left panel) and also ablated neurons in the olfactory bulb bilaterally as a control (Figure 2A right panel) in zebrafish larvae of a Gal4 line gSAIzGFFM119B that were mated with UAS:EGFP reporter fish. Results show that two-photon laser can ablate targeted cells while leaving adjacent cells or neurites unaffected (Figure 2B).
The pretectal neurons project their axons toward the ILH ipsilaterally. Here, we investigated their functional connectivity using Ca imaging. Neuronal activity in this region can be observed as Ca signals with a Ca probe GCaMP6s (Figure 4A), whose expression was driven by a Gal4 line gSAIzGFFM119B (Figure 4B) or another Gal4 line hspGFFDMC76A (Figure 4C). The colored arrowheads (Figure 4) show swimming directions and trajectories of the paramecium, as well as color-coded Ca signal-changes in the specified brain area, which were evoked by the sight of the swimming paramecium (Figure 4B-D). Both the pretectum (Figure 4B) and the ILH (Figure 4C) showed neuronal activity in the proximal presence of prey, suggesting that both neuronal activities are visually driven.
By using the double Gal4 GCaMP6s larvae (gSAIzGFFM119B; hspGFFDMC76A; UAShspzGCaMP6s), we ablated the pretectal neurons unilaterally and further performed Ca imaging to observe neuronal activity in the pretectum and the ILH in the ablated larvae. The left pretectum that was laser-ablated showed residual to no neuronal activity (Figure 4D, top left), suggesting that laser-ablation was successful. However, the ipsilateral ILH showed dramatically reduced neuronal activity (Figure 4D bottom left), suggesting that the major input to the ILH comes from the ipsilateral pretectum. In contrast, the ILH on the other side of the laser-ablated pretectum (Figure 4D bottom right) showed neuronal activity comparable to the neuronal activity in the ipsilateral pretectum (Figure 4D, top right), which suggests that the right ILH is receiving inputs from the right pretectum.
The choice of which behavioral assay to use in an experiment depends on the possible role(s) of the neurons under investigation. Here, we show an example of a prey-capture assay following ablation of a pretectal nucleus that was identified as a prey detector21. Using the lighting system shown in Figure 5A-D, the number of paramecium in the recording chamber can be counted by image processing (Figure 5E-G). Automated extraction of eyes, and calculation of eye positions (i.e., changes in the angles of the eyes) can also be possible because the eyes of the zebrafish larva appear darker than the background in this lighting condition (Figure 5H).
Results of the experiment show that bilateral-ablation of the pretectum abolished prey-capture activity (Fig. 5I, PT) while ablation of a subpopulation of neurons in the olfactory bulb did not (Figure 5I, OB). These results, together with experiments shown in Figure 4, suggest that the pretecto-hypothalamic circuit is essential in prey-capture behavior.
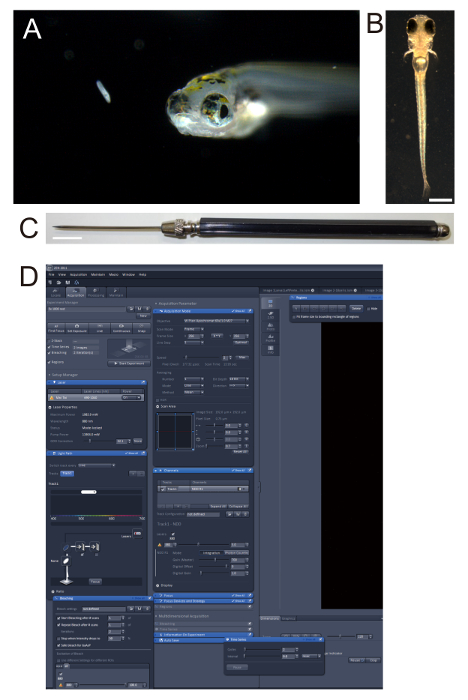
Figure 1. Zebrafish larvae. (A) Five-day old (5-day post-fertilization (5-dpf)) zebrafish larva and its prey, a paramecium. (B) 4-dpf nacre larva embedded in 2% low melting-point agarose. Note that the nacre strain lacks black pigments on the body while the retinal pigment epithelium is intact. Scale bar: 0.5 mm. (C) Dissecting needle used for orienting zebrafish larvae in agarose. Scale bar: 1 cm. (D) Screenshot of the software used in the two-photon microscopy, showing "Bleaching", "Time Series", and "Regions" panels that were used in ablation. Please click here to view a larger version of this figure.
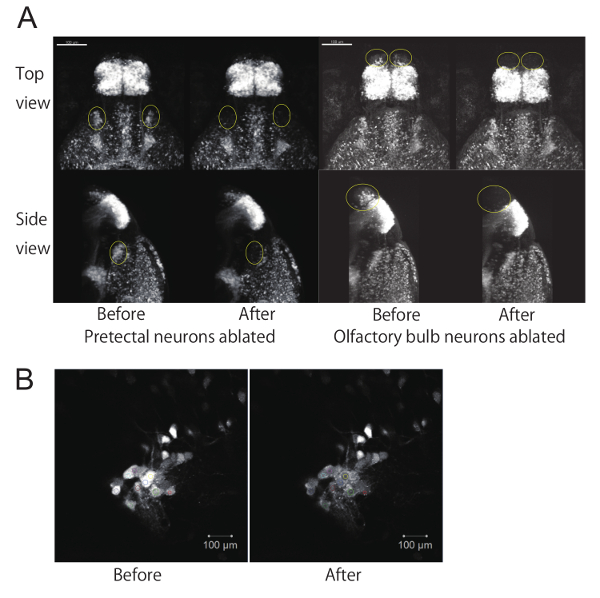
Figure 2. Ablation of a subpopulation of neurons using a two-photon laser microscope. (A) EGFP fluorescent images of 4-dpf zebrafish larvae before and after ablation of neurons. Top view and side view of 3D reconstructed z-stack images using image processing software. The z-stack images were taken with a 20X objective lens. The ablated areas (either the pretectum or olfactory bulb) are circled in yellow. Scale bar: 100 µm. (B) Example of laser ablation at a single focal plane using the "Bleaching" function. Laser-irradiated areas (set as ROIs) are shown in colors on each cell. Scale bar: 10 µm. Please click here to view a larger version of this figure.

Figure 3. Preparation of zebrafish larvae and paramecia for Ca imaging. (A) Recording chamber. A commercially available 9 mm diameter x 0.8 mm depth hybridization gasket with 8 chambers was utilized as the recording chamber. The gasket was put on a glass slide and adhered. The seal on the top of the gasket was peeled off. (B) The nylon mesh (32 µm) used to rinse the paramecia. The mesh was placed on a 50-mL tube. (C) Paramecium culture containing rice straw and dry yeast pellets. (D) Paramecium stock solution prepared from the culture shown in C. (E) Paramecium stock solution was rinsed with system water twice to remove possible olfactory and gustatory cues in the medium. (F) Paramecium embedded in the recording chamber. To remove a portion of agarose, several cuts were made. (G) Recording chamber with a zebrafish larva and a paramecium. The majority of the agarose was removed to allow the paramecium to swim. The head of the zebrafish larva is exposed. (H) Upright fluorescent microscope used in Ca imaging. The microscope is equipped with a scientific CMOS camera. (I) Zebrafish larva in the recording chamber under the microscope with the excitation light on. With a 2.5X objective lens, the illuminated area is slightly larger than the size of the chamber. Please click here to view a larger version of this figure.
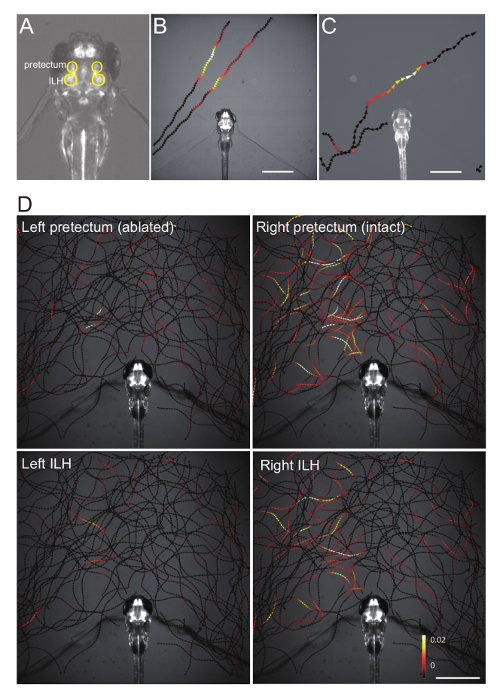
Figure 4. Representative Ca imaging data. (A) Position of the pretectum and the inferior lobe of the hypothalamus (ILH), circled in yellow, in a double Gal4 hspGFFDMC76A; gSAIzGFFM119B; UAShspzGCaMP6s zebrafish larva. (B) Changes in the pretectum Ca signal (averaged bilaterally), color-mapped onto the trajectories of a paramecium in a gSAIzGFFM119B; UAShspzGCaMP6s larva. Scale bar: 1 mm. (C) Changes in the ILH Ca signal (averaged bilaterally), color-mapped on the trajectories of a paramecium in a hspGFFDMC76A; UAShspzGCaMP6s larva. Scale bar: 1 mm. (D) Changes in the pretectum and the ILH Ca signals color-mapped on the trajectories of a paramecium in a hspGFFDMC76A; gSAIzGFFM119B; UAShspzGCaMP6s larva that was subjected to two-photon laser-ablation of the left pretectum. Scale bar: 1 mm. This image was reproduced from the same data previously published21. Please click here to view a larger version of this figure.

Figure 5. Prey capture assay and representative data in laser-ablated zebrafish larvae. (A) Behavioral recording system. The stereomicroscope was equipped with a CMOS camera. Images were acquired using a custom-made script. (B) Components of the lighting system. A grey-colored paper, a glass diffuser, white LED ring light, diffuser, a custom-made spacer (cardboard), and diffuser with a hole at the center. (C) The lighting system assembled from the parts shown in B. (D) Recording chamber for the prey capture assay. A chamber (diameter: 20 mm; depth: 2.5 mm) was placed on a glass slide. The original top seal of the chamber was peeled off. A glass-cover was put on the recording chamber. (E) Raw image from a frame of the recorded movie. (F) A frame divided (pixel-by-pixel) by an average image in a movie. Note that the non-uniform background in lighting can be compensated by this image processing. (G) Paramecia extracted from a frame using the "Particle Analysis" function in Fiji. (H) Example of image in Fiji with an applied threshold. Paramecia appear bright, whereas the eyes of the zebrafish larva appear dark, and the background is grey. Changes in eye positions (i.e., angles with respect to the body axis) can be calculated, if necessary. (I) Representative experimental data of paramecium consumption in an olfactory-bulb-ablated control larva (OB) and a pretectum-ablated larva (PT). Pretectum ablation significantly reduced prey hunting ability while olfactory bulb ablation did not affect it. Please click here to view a larger version of this figure.
