Through the injection of the pathogen directly in the deep derma, the structural morphology of the tissue remains intact (Figure 1A).
The maintenance of skin structural integrity allows the detection of immune cells and their localization at the site of infection. The higher magnification shown in Figure 1B reveals that the abscess is mainly composed of polymorphonuclear leukocytes (PMCs) that contain the spread of the pathogen by its confinement to the site of infection. Recruitment of PMCs, such as neutrophils, allows the formation of the abscess, thus avoiding the spreading of the fungus within the tissue14. Higher magnifications (Figure 1C, D) show how the surrounding areas of the abscess are also enriched in PMCs.
By maintaining the structural integrity, the localization of the pathogen during the infection can be determined. Taking advantage of the elevated content of polysaccharides in the cell wall of C. albicans, the identification of the pathogen was performed using PAS staining, which gives a purple-magenta color to the fungus. As shown in Figure 2A, PAS staining clearly shows that the pathogen is confined inside the abscess, formed by the recruitment of granulocytes14. C. albicans is clearly visible at the higher magnification (Figure 2B), and it appears to be surrounded by necrotic and immune cells.
Using the method described above, the infection process and the consequent inflammation can be followed over time. At the very beginning, immune cell recruitment leads to the formation of an abscess (Figure 1A and Figure 3A). When the analysis of the skin tissue was prolonged up to 48 – 72 h, the disruption of the skin structures and the formation of a scar was visualized (Figure 3B, C). The formation of this structure is due to the healing phase that follows the expulsion of the pathogen14.
The process that leads to the formation of an ulcer can be followed for several days after the infection. As shown in Figure 4A, C57BL/6 wild-type mice display an ulcerative process that increases over time. Around 70 – 80% of the mice show formation of an ulcer 72 h after infection. Due to experimental variability, at least 8 mice should be used in every experiment. For statistical analysis, it is recommended to use a minimum of 8 – 10 mice for each ulceration score if comparing different genotypes or treatments. In this case, even small differences should be visible. Illustrative pictures of the ulcerative process are shown in Figure 4B. Upon specific treatments or genetic deficiencies, the formation of the ulcer can be reduced and/or abrogated14. In some cases, instead of the ulcer, a cyst is formed, as illustrated in Figure 4C.
Other than Hematoxylin & Eosin and PAS staining, the slides can be stained for either specific cellular markers or with other immunohistochemical stainings to better visualize: collagen fibers; cytokine/chemokines location within the tissue; and immune cell identity and spatial distribution.
In addition to histological preparation, samples can be used also for molecular analyses. The entire injection site can be excised, as for histological preparations, and processed for RNA extraction. RNA extraction from the tissue allows the study and identification of those genes upregulated during the infection with the pathogen. The possibility to process the samples for molecular analyses is an important tool, together with histological analysis, in order to better characterize the type of immune response elicited against the pathogen and its regulation during the early and late stages after the challenge14.
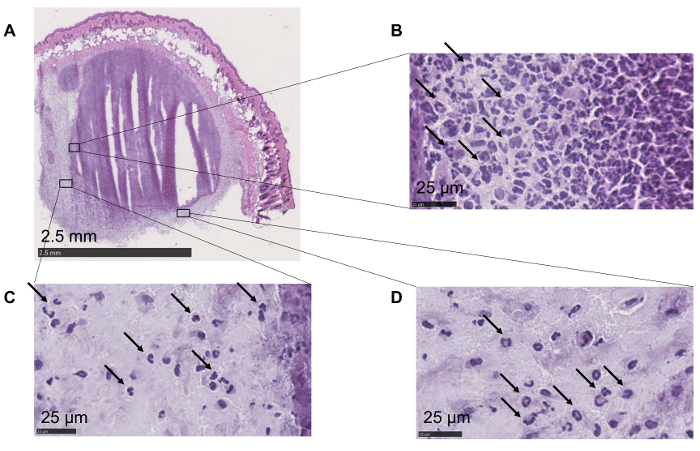
Figure 1: Maintenance of the skin morphology and cellular localization. (A) All the layers are preserved. Epithelium, adipocytes, and muscle cells are easily recognizable. (B) Polymorphonuclear leukocytes (highlighted with arrows) can be identified within the abscess. (C, D) Higher magnifications show the presence of polymorphonuclear leukocytes (arrows) in the area surrounding the abscess. Please click here to view a larger version of this figure.
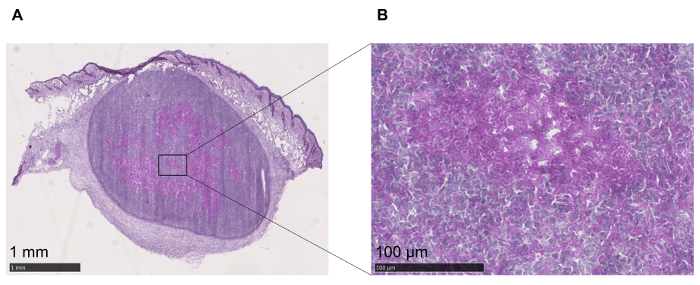
Figure 2: Identification of C. albicans. (A) C. albicans can be detected within the tissue with PAS staining. (B) Higher magnification to better visualize the fungus, stained in purple, confined inside the abscess. Please click here to view a larger version of this figure.
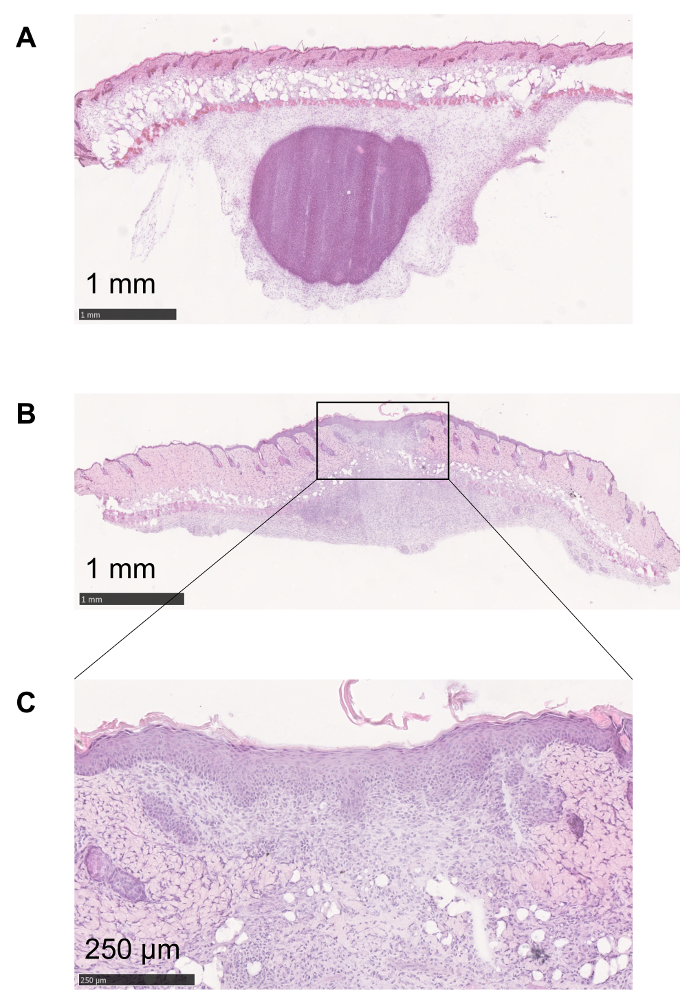
Figure 3: Follow up of the inflammatory process. (A) At the very beginning of the inflammatory process elicited by C. albicans injection, granulocytes are recruited in order to form an abscess to confine the pathogen. (B, C) The formation of an ulcer is necessary for the expulsion of the fungus. The healing tissue is characterized by an alteration in the structural integrity of the skin and can lead to the formation of a scar. Please click here to view a larger version of this figure.
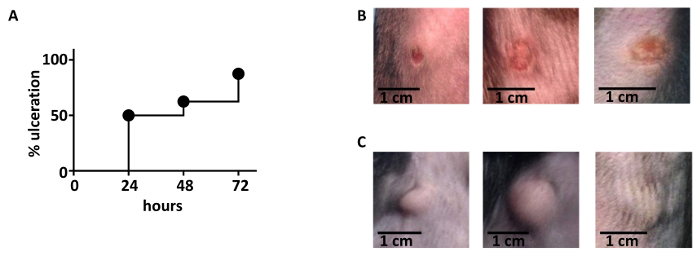
Figure 4: Kinetics of ulceration. (A) The ulcerative process follows a trend of increase during the first days after the infection. 72 h after the infection, 70 – 80% of the mice shows an ulcer at the site of infection. 16 C57BL/6 wild type animals were used. (B) Illustrative pictures of ulcers that developed 48 h after infection. (C) Illustrative pictures of cyst formation after infection. Please click here to view a larger version of this figure.
