The results of repeated evaluations of respiratory function by DCP, performed under baseline conditions on three consecutive days (days 12, 13, and 14 of the protocol illustrated in Figure 1) in control and allergic BALB/c mice, are shown in Figure 3. The parameters that were selected to assess the pattern of breathing included breathing frequency (Figure 3A), tidal volume (Figure 3B), minute ventilation (Figure 3C), and end-inspiratory pause (Figure 3D). The parameters used to assess airway obstruction were EF50 (Figure 3E) and sRaw (Figure 3F). The outcomes of each selected parameter were stable over these three consecutive days in both groups, with no apparent effect caused by allergic inflammation.
The degree of responsiveness to methacholine was evaluated by DCP on successive days (days 12, 13, and 14 of the protocol illustrated in Figure 1) in both control and allergic BALB/c mice. The results, shown in Figure 4, display the changes in the two parameters that are sensitive to airway obstruction, namely sRaw (Figure 4A, B and C) and EF50 (Figure 4D, E and F). As expected, incremental concentrations of methacholine progressively increased sRaw and progressively decreased EF50. These responses were potentiated by allergic inflammation, especially at the final concentration tested, which attested the presence of hyperresponsiveness. The results also demonstrate that the exaggerated degree of responsiveness was confined to the first day (day 12), as it was not observed during the two subsequent assessments (i.e., at days 13 and 14).
The results of the assessment of respiratory mechanics by the FOT, performed on the last day of the experimental protocol (day 15; Figure 1) in both control and allergic BALB/c mice, are shown in Figure 5. These experiments were included in the study to complement the DCP evaluations. The FOT is recognized as a more precise approach to assess respiratory function2. One of its strengths is that it provides topographic insights as to which sites of the lung (conducting airways versus peripheral airways and lung tissue) are affected by the tested interventions (e.g., allergen and methacholine). The recommended methodology to assess respiratory mechanics with the FOT was previously described1. Herein, three FOT parameters were used to describe the changes in respiratory mechanics induced by allergic inflammation and methacholine. These parameters included: 1-Newtonian resistance (RN; Figure 5A), a parameter for which the changes in value mainly reflect variations in the resistance of the large conducting airways; 2-tissue damping (G; Figure 5B), a parameter for which the changes in value mainly reflect variations in tissue resistance; and 3-tissue elastance (H; Figure 5C), a parameter for which the changes in value mainly reflect variations in tissue stiffness2. As expected, there was an increase in each of these parameters in response to incremental concentrations of methacholine. Consistent with sRaw and EF50 results obtained with the DCP from the previous day (day 14; Figure 1), the changes in RN induced by methacholine (Figure 5A) were comparable between the control and the allergic mice. In fact, the values of sRaw at day 14 correlated with the values of RN at day 15 (Figure 5D). The rise in H induced by methacholine was also similar between the control and the allergic mice (Figure 5B). However, the methacholine-induced increase in G was significantly greater in the allergic mice (Figure 5C). This result demonstrates the presence of a persisting hyperresponsive phenotype in the allergic mice at day 15, which was not detected by the DCP evaluations performed on the two previous days.
The entire study was repeated with C57BL/6 mice. The results of the successive DCP evaluations of sRaw, at days 12, 13, and 14 of the protocol (Figure 1), and of the FOT evaluation of RN, at day 15, are shown in Figure 6. In that specific mouse strain, the exaggerated methacholine response observed in the allergic mice was maintained over the three consecutive days (Figure 6A, B, and C). This hyperresponsive phenotype was also depicted with the FOT at day 15 by a rise in RN induced by methacholine that was more pronounced in the allergic mice (Figure 6E). These were in stark contrast with the results obtained with the BALB/c mice, where a progressive waning of the hyperresponsiveness occurred from days 12 to 14 (Figure 4) and a lack of difference in the methacholine-induced rise in RN was observed at day 15 (Figure 5A). Together, these results indicated a time-varying effect of the allergen on the methacholine-induced response between the two strains of mice. Importantly, this strain difference was depicted by both the DCP and the FOT. Concordantly, the values of sRaw measured by the DCP at day 14 correlated with the values of RN measured by FOT at day 15 (Figure 6F), as was observed with the BALB/c mice (Figure 5D).
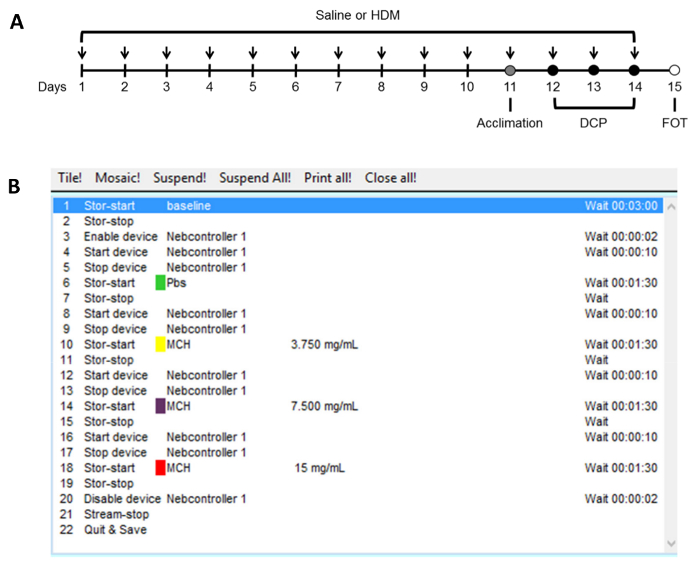
Figure 1. Protocols used to induce pulmonary allergic inflammation and to assess the degree of responsiveness to methacholine. This study was conducted on female BALB/c and C57BL/6 mice of 7 to 9 weeks of age. The sequence of interventions carried out over the entire study is shown in panel (A). Half of the mice was exposed to 50 µg of house-dust mite (HDM) extract intranasally on 14 consecutive days to induce pulmonary allergic inflammation. The other half was exposed to saline and used as control. Respiratory function was assessed by double-chamber plethysmography (DCP) on three separate occasions (Days 12, 13, and 14; black circles) following an acclimation session (Day 11; gray circle) that included a challenge with aerosolized saline. During each session, baseline respiratory function and the response to methacholine were assessed using the automated protocol shown in panel (B). At day 15, an invasive assessment of respiratory mechanics with the forced oscillation technique (FOT) was performed as previously described1. Please click here to view a larger version of this figure.
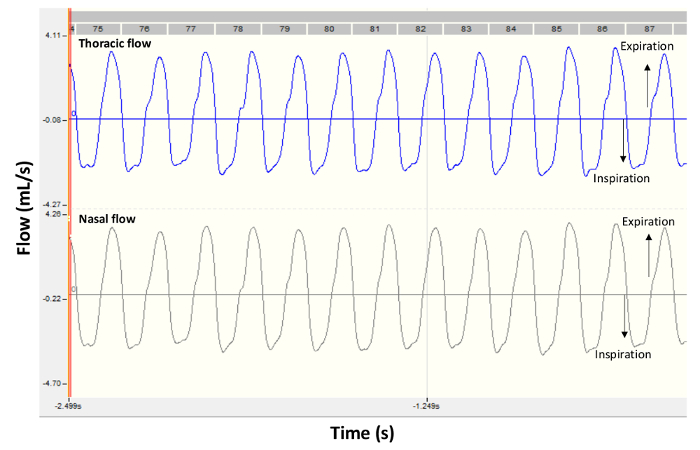
Figure 2. Representative flow signals from a healthy BALB/c mouse. The panels show typical recording traces obtained by double-chamber plethysmography in a control mouse under baseline conditions. Thoracic flow is shown in the upper panel and nasal flow is shown in the lower panel. Negative values are during inspiration and positive values are during expiration. Please click here to view a larger version of this figure.
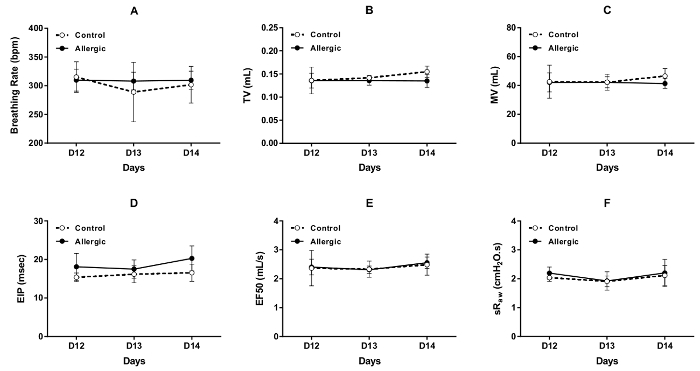
Figure 3. Repeated evaluations of respiratory function in conscious BALB/c mice. Baseline respiratory function was assessed by double-chamber plethysmography (DCP) in control (open symbols) and allergic (solid symbols) mice on days 12, 13, and 14 of the protocol illustrated in Figure 1. The DCP parameters used to assess respiratory function included breathing frequency in (A), tidal volume (TV) in (B), minute ventilation (MV) in (C), end inspiratory pause (EIP) in (D), flow at mid-tidal expiratory volume (EF50) in (E), and specific airway resistance (sRaw) in (F). The values of breathing frequency, TV, MV, sRaw and EIP for each mouse were the average values recorded over 1.5 min. The value of EF50 was the minimal value obtained during this recording period. The results are presented as group means ± standard deviation (n=5/group). Please click here to view a larger version of this figure.
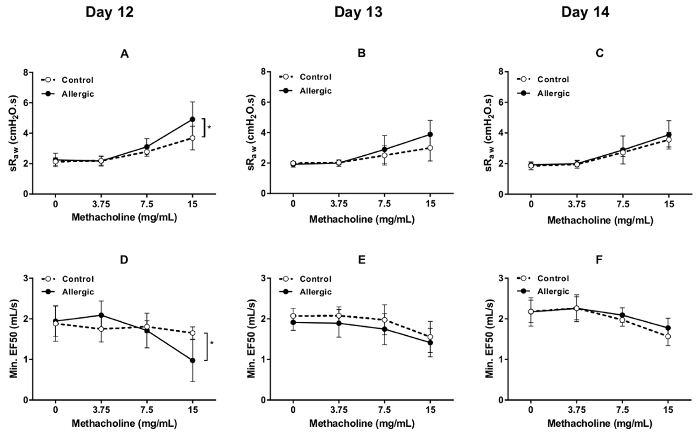
Figure 4. Methacholine bronchoprovocation test in conscious BALB/c mice. Methacholine responsiveness was assessed by double-chamber plethysmography (DCP) in control (open symbols) and allergic (solid symbols) mice on days 12, 13, and 14 of the protocol illustrated in Figure 1. The DCP parameters used to assess the response included specific airway resistance (sRaw) in (A) through (C) and flow at mid-tidal expiratory volume (EF50) in (D) through (F). The bronchoprovocation was performed by aerosolizing methacholine in the DCP head chamber for 10 s at incremental concentrations. The response was monitored during 1.5 min after each concentration. The value of sRaw for each mouse at each concentration was the average value recorded over 1.5 min. The value of EF50 was the minimal value obtained during this recording period. The results are presented as group means ± standard deviation (n=5/group). The asterisk symbol * designates a statistically significant difference (p 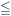 0.05). Please click here to view a larger version of this figure.
0.05). Please click here to view a larger version of this figure.
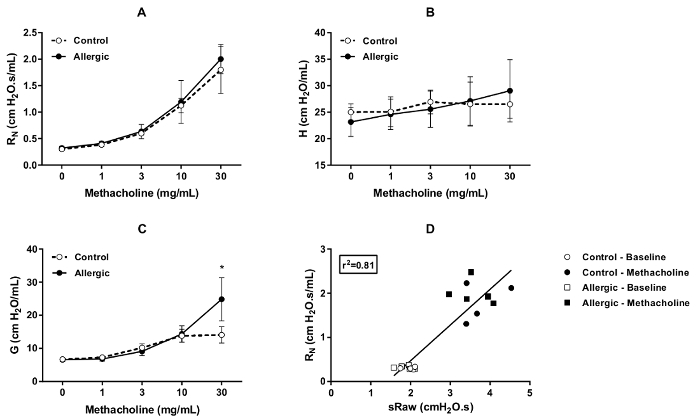
Figure 5. Invasive assessment of respiratory mechanics in BALB/c mice. Respiratory mechanics at baseline and in response to methacholine was evaluated by the forced oscillation technique (FOT) on Day 15 of the protocol illustrated in Figure 1. The control (open symbols) and the allergic (solid symbols) mice were the same as the ones tested by double-chamber plethysmography (DCP) on Days 12, 13, and 14. The parameters used to assess respiratory mechanics were Newtonian resistance (RN) in (A), tissue elastance (H) in (B) and tissue damping (G) in (C). The bronchoprovocation was performed by nebulizing incremental concentrations of methacholine directly into the endotracheal tube of anesthetized, tracheotomized, paralyzed, and mechanically ventilated mice in supine position. The response was monitored during 5 min after each concentration. The value for each parameter for each mouse at each concentration was the peak value obtained during this recording period. The results are presented as group means ± standard deviation (n=5/group). Panel (D) shows the correlation between specific airway resistance (sRaw) measured by DCP on day 14 and RN measured by FOT on day 15. The open symbols represent the values at baseline and the solid symbols represent the maximal values at the highest concentration of methacholine tested for either the control (circles) or the allergic (squares) mice. The inset shows the coefficient of determination (r2). The asterisk symbol * designates a statistically significant difference (p ≤ 0.05). Please click here to view a larger version of this figure.
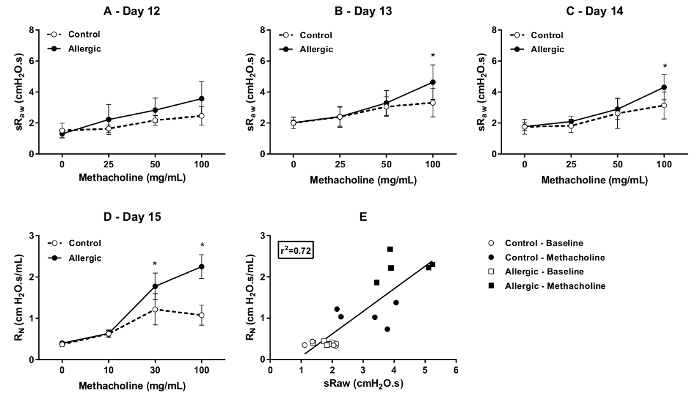
Figure 6. Respiratory function and invasive respiratory mechanics in C57BL/6 mice. Specific airway resistance (sRaw) was assessed by double-chamber plethysmography (DCP) at baseline and in response to methacholine in control (open symbols) and allergic (solid symbols) mice on days 12 (A), 13 (B), and 14 (C) of the protocol illustrated in Figure 1. Newtonian resistance (RN) at baseline and in response to methacholine were assessed by the forced oscillation technique (FOT) on day 15 (D). The bronchoprovocations were performed as described in Figure 4 and Figure 5 for the DCP and the FOT, respectively. The results are presented as group means ± standard deviation (n=5/group). Panel (E) shows the correlation between sRaw measured by DCP on day 14 and RN measured by FOT on day 15. The open symbols represent the values at baseline and the solid symbols represent the maximal values at the highest concentration of methacholine tested for either the control (circles) or the allergic (squares) mice. The inset shows the coefficient of determination (r2). The asterisk symbol * designates a statistically significant difference (p ≤ 0.05). Please click here to view a larger version of this figure.
| Parameter | Unit | Description | Information |
| F | bpm | Breathing frequency | Pattern of ventilation |
| TV | mL | Tidal volume | |
| MV | mL | Minute ventilation | |
| Ti | ms | Inspiratory time | |
| Te | ms | Expiratory time | |
| PIF | mL/s | Peak inspiratory flow | |
| PEF | mL/s | Peak expiratory flow | |
| EV | mL | Expiratory volume | |
| NTV | mL | Nasal tidal volume | |
| NEV | mL | Nasal expiratory volume | |
| EIP | ms | End inspiratory pause | |
| EEP | ms | End expiratory pause | |
| dT | ms | Time delay | Airflow obstruction |
| sRaw | cmH2O·s | Specific airway resistance | |
| sGaw | 1/cmH2O·s | Specific airway conductance | |
| EF50 | mL/s | Flow at mid-tidal expiratory volume | |
| Sr | % | Success rate | Quality control |
| N | Number of valid breaths |
Table 1. List of typical parameters obtained from double-chamber plethysmography. The parameters were grouped according to the nature of the information they provide during a respiratory function evaluation.
| Advantages | Limitations |
| · Conscious animals | · Necessity to control the surrounding environment |
| · Accurate ventilation parameters | · Prior acclimation of the animals |
| · Undisputed indexes of airflow obstruction (sRaw, EF50) | · Requirement to hermetically separate the nasal and thoracic flows |
| · Adaptable to various species and animal sizes | · Absolute value variability for some outcome parameters |
| · Used in many research applications | · sRaw not a true measurement of resistance |
| · Straightforward technique | · Presence of the upper airways |
| · Sensitive to change | · Complementing the measurements with an invasive assessment |
Table 2. List of advantages and limitations associated with double-chamber plethysmography.
| Double-Chamber Plethysmography | Forced Oscillation Technique | |
| Animal’s state of consciousness | Unaltered | Anesthetized (and usually paralyzed) |
| Animal’s position | Upright | Supine |
| Animal’s accessibility | Confined within chamber | Accessible |
| Animal integration to the measurement device | Nose or neck seal | Tracheotomy or oral intubation |
| Animal’s airway tree | Intact | Partial – upper airway segment excluded (i.e. nasal conducts, pharynx and larynx) |
| Lung volume at which the outcome parameters are obtained | Variable – spontaneous volume adopted by the animal | Standardized – using controlled recruitment maneuvers and positive end-expiratory pressure. |
| Frequency at which the outcome parameters are assessed | Variable – spontaneous breathing frequency adopted by the animal | Controlled – using predefined waveforms at specified frequencies |
| Contribution from upper airway segment to the outcome parameters | To be expected | Circumvented |
| Site of aerosol delivery | Inside the head chamber | Directly in the trachea |
| Effect of upper airway segment on inhaled dose/ aerosol deposition pattern | To be expected | Prevented |
| Ability to detect change – based on the results of the present study | Observed | Observed |
| Inherent variability of the technique – based on the results of the present study | Fluctuation of the coefficient of variation for sRaw at baseline: 7.5 – 20.6% | Fluctuation of the coefficient of variation for RN at baseline: 3.6 – 13.4% |
Table 3. Comparison between the double-chamber plethysmography and forced oscillation techniques.
