The in situ hybridization assay workflow:
The workflow is depicted in Figure 1 and consists of four parts: permeabilization of cells or tissues with target retrieval and protease solutions, hybridization of the probes to the target RNA, signal amplification, and visualization of the signal. The signal can also be quantified using digital imaging software systems or in a semi-quantitative manner based on the number of dots per cell. The manual procedure described in Figure 2 has also been fully automated in commercial auto-staining systems.
Representative staining for exon junction detection (EGFRvIII splice variant): EGFRvIII is a variant of the epidermal growth factor receptor that arises from an in-frame genomic deletion of exons 2 to 7, leading to constitutively active oncogenic signaling7. The assay was used to identify EGFRvIII status in FFPE glioblastoma (GBM) tumor samples. Single double-Z probes were designed to span the exon junctions in order to detect either WT, mutant, or both transcripts (Figure 3A). The WT EGFR probes span the junctions of either exons 1 and 2 (E1/E2) or exons 7 and 8 (E7/E8), while the EGFRvIII-specific probes span the junction of exons 1 and 8 (E1/E8). A common probe that spans the junction of exons 8 and 9 (E8/E9) was also used to detect total EGFR (both WT and EGFRvIII transcripts). All the probes were then used to determine EGFR status in FFPE GBM samples. The two representative examples shown in Figure 3B were taken from a larger study. EGFR status was confirmed by an independent method, RT-PCR. Both WT probes detected signal in both samples, indicating that the two samples express WT EGFR (Figure 3B). However, the mutant probe only showed signal detection in the EGFRvIII+ sample, confirming that this sample is indeed positive for the EGFRvIII variant (Figure 3B, left panels). Conversely, the mutant probe did not detect signal in the EGFRvIII- sample (Figure 3B, right panels). Taken together, these results demonstrate that the exon junction assay can identify EGFRvIII status in GBM FFPE tumor samples.
Representative staining for short targets:
The CDR3, or complementary determining region 3, is a highly variable domain in T-cell receptors. Typically, the CDR3 sequence are quite short; for example, the CDR3 α and β sequences from the Jurkat T-cell line are 51 and 48 nucleotides in length, respectively (Figure 4A). To identify the specific CDR sequences expressed in Jurkat cells, antisense probes for CDR3 α and β that are expressed in the Jurkat T-cell line were generated, in addition to sense probes for CDR3 α and β to serve as negative control probes. All the probes were then tested in FFPE-prepared Jurkat cells with the assay. Robust staining was observed with anti-sense probes for both CDR3 α and β in the Jurkat cells, whereas sense probes detected little to no signal (Figure 4B). These results demonstrate the ability of the short target assay to discern between highly variable but short CDR sequences for T-cell receptor clones.
Representative staining for point mutation EGFR L858R:
Point mutation probes were developed to detect single nucleotide variations and small insertions or deletions (INDELs) in the tumor context. Figure 5A demonstrates the ability for in situ detection of the point mutation EGFR L858R (2573T>G). Two probes were designed: one to detect the L858R mutated EGFR sequence, and another to detect the EGFR L858 WT sequence. Both probes were tested in two FFPE-prepared cell lines: H2229, which only expresses EGFR L858 WT; and H1975, which is heterozygous for the EGFR L858R mutation. The L858R mutant probe detected signal only in the H1975 cell line but not in the H2229 cell line. However, the WT probe detected signal in both cell lines. Similarly, Figure 5B visualizes in situ detection of the point mutation KRAS G12A (35 G>C). Two probes were designed to detect the KRAS G12A MT and KRAS G12 WT sequences and then tested on the HuT78 cell line (which only expresses KRAS G12 WT) and the SW116 cell line (which is heterozygous for the KRAS G12A mutation). While the KRAS G12 WT probe detected signal in both cell lines, the KRAS G12A probe only detected signal in the SW116 cell line. Taken together, these data demonstrate the technical capability of the point mutation assay in detecting single nucleotide polymorphisms in the cell and tissue context.
Representative staining for the automated in situ assay:
Automated assays allow for a greater number of samples to be run more reliably, minimizing inter-user variability and hands-on time and generating consistently reproducible results. Therefore, an automated version of the assay was developed. To demonstrate automated staining with this assay, detection of the splice variant METΔ14 was examined. This variant is the result of exon 14 in the MET gene being skipped during pre-mRNA splicing, which leads to constitutive activation and oncogenic transformation of the MET receptor8,9. To specifically detect the METΔ14 variant, two exon junction probes were designed: one that spans the junction of exons 13 and 15 (E13/E15) to detect the METΔ14 variant transcript, and another that spans the junction of exons 14 and 15 (E14/E15) to detect the WT MET transcript (Figure 6A). Both probes were then tested in 2 FFPE-prepared cell lines: H596, which expresses the METΔ14 variant, and A549, which expresses the WT MET gene. Both probes showed mutually exclusive expression patterns, with the E13/E15 probe only detecting signal in the H596 cells and the E14/E15 probe only detecting signal in the A549 cells (Figures 6B and 6C). Lastly, the probe for dapB showed no signal, indicating no background signal (Figures 6B and 6C). Overall, this data demonstrates specific detection of the MET variant METΔ14 in situ utilizing the automated BaseScope assay.
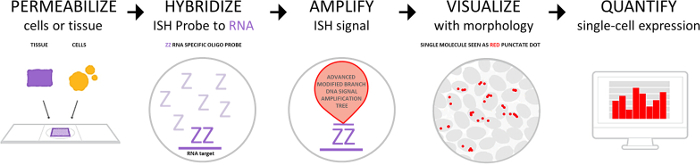
Figure 1: The assay workflow. The workflow consists of 4 major steps: pretreatment to permeabilize cells or tissue, probe hybridization to target RNA, signal amplification, and signal detection by visualization under brightfield or fluorescent microscope. Individual dots can be quantified using a digital image analysis platform. Please click here to view a larger version of this figure.
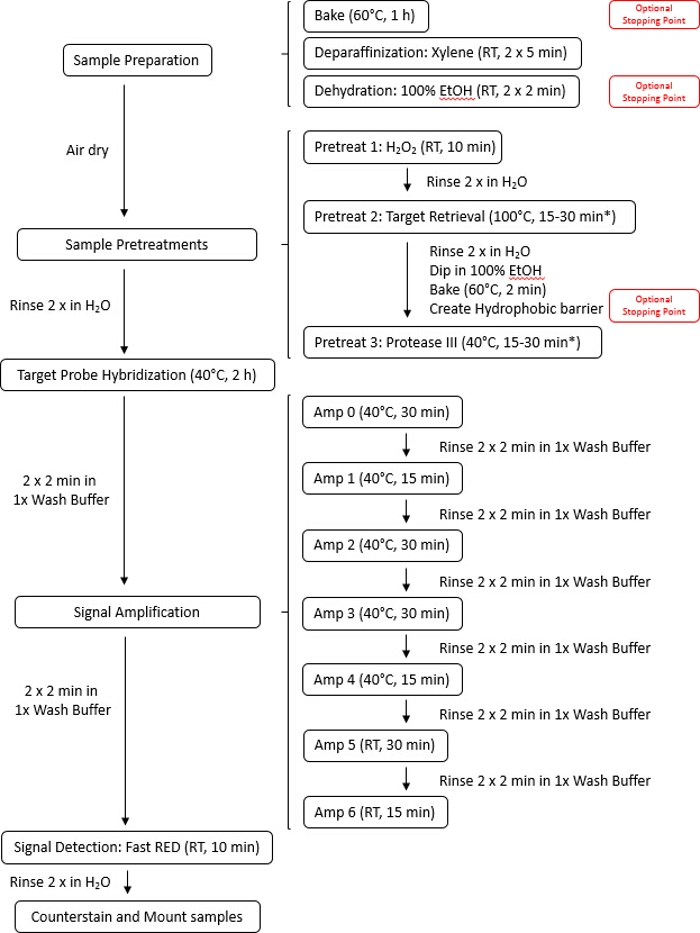
Figure 2: Illustration of the manual assay protocol. The entire assay can be completed in 9 h. Pretreatment times may vary depending on tissue type, so it is advised to consult Appendix A in the user manual for tissue pretreatment recommendations regarding incubation time. Please click here to view a larger version of this figure.
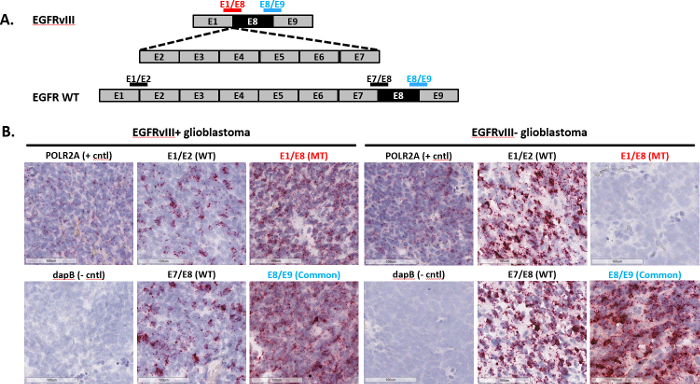
Figure 3: Representative images of exon junction detection. (A) Shown here are the exon organization for EGFR WT and EGFRvIII transcripts and a schematic depicting double-Z exon junction probes that straddle the junction to detect EGFR WT or EGFRvIII. (B) The exon junction assay was performed on two FFPE glioblastoma samples using the probes listed in (A), as well as a positive control probe, POLR2A, and negative control probe, dapB. Please click here to view a larger version of this figure.
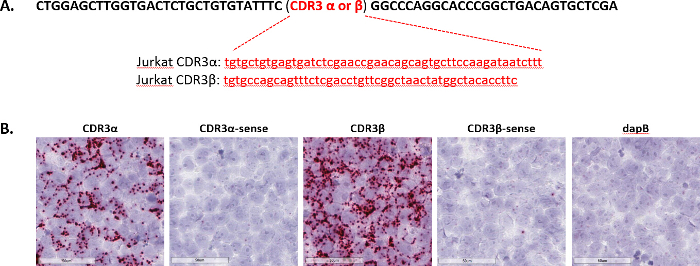
Figure 4: Representative images of short target sequence detection. (A) Shown here are CDR3 sequences in Jurkat cells. The sequence in black is a flanking common sequence and the sequence in red is unique to either CDR3α or CDR3β. Single double-Z short target sequence probes were designed against these sequences. (B) The short target assay was performed on Jurkat cells prepared as an FFPE cell pellet using anti-sense or sense probes targeting the sequences in (A), and dapB was used as a negative control. Please click here to view a larger version of this figure.
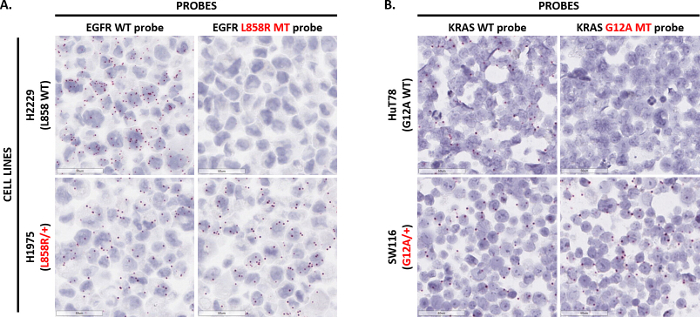
Figure 5: Representative images of point mutation detection. (A) The assay was performed on H2229 cells (homozygous for EGFR L858) and H1975 cells (heterozygous for the EGFR L858R mutation) prepared as an FFPE cell pellet using single double-Z point mutation probes targeting the EGFR L858 WT sequence or EGFR L858R mutated sequence. (B) The point mutation assay was performed on HuT78 cells (homozygous for KRAS G12A) and SW116 cells (heterozygous for the KRAS G12A mutation) prepared as an FFPE cell pellet using single double-Z point mutation probes targeting the KRAS G12 WT sequence or KRAS G12A mutated sequence. Please click here to view a larger version of this figure.
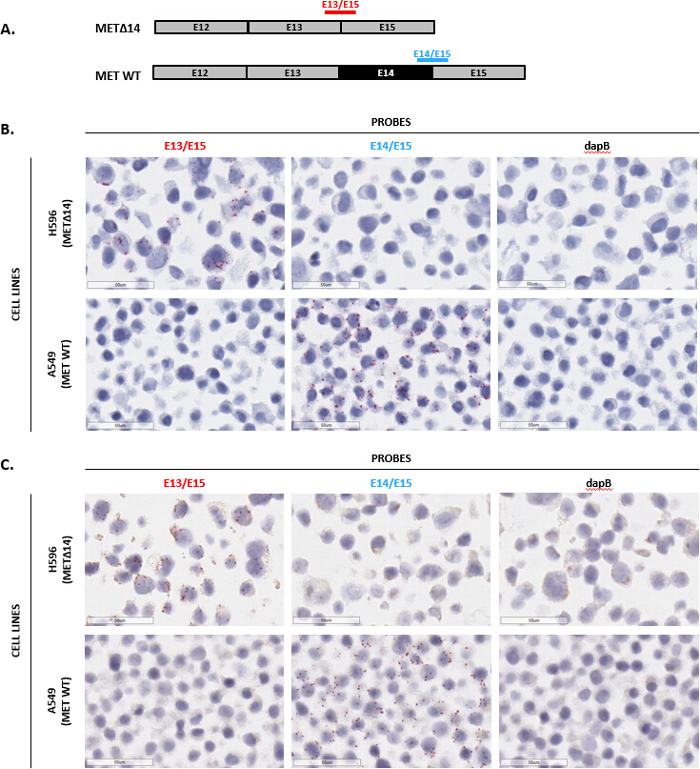
Figure 6: Representative images of automated staining with the exon junction assay. (A) Shown here is the exon organization for MET WT and METΔ14 transcripts and a schematic depicting single double-Z exon junction probes that straddle the junction to detect MET WT or METΔ14. (B) and (C) The automated exon junction assay was performed on H596 cells (expressing METΔ14) and A549 cells (expressing MET WT) prepared as an FFPE cell pellet using the probes listed in (A), as well as negative control probe dapB. Please click here to view a larger version of this figure.
| Exon Junction | Short Sequence | Point Mutation |
| Splice variant/isoform | Sequences between 50 and 300 nt | Point mutation |
| Circular RNA (circRNA) | Highly homologous sequences | Short indel |
| Gene fusion | CDR3 sequence for TCR clones | Gene editing |
| Gene knockout (KO) | Pre-miRNA | |
| Small nucleolar RNA (snoRNA) | ||
| Gene editing |
Table 1: Applications of the in situ assay. There are 3 main categories for applications of this assay: exon junction, short target sequence, and point mutation. Listed in each column are some specific application examples for each category.
