Positive results showing RCR and proton leak kinetics are shown in Table 1 and Figure 15, respectively. In this study7, RCR and protein leak kinetics were measured in Holstein dairy cows at 70 days in milk after cows had been fed 1 of 5 different levels of Cu, Zn and Mn for 28 days. State 4, maximum proton leak-dependent respiration, had a tendency to be affected by mineral intake of Cu, Mn and Zn (p < 0.1). State 3 respiration (maximum ATP stimulated respiration) and RCR = State 3 / State 4 (respiratory control ratio) was not affected by mineral intake. State 4 respiration was highest in LowMn and lowest in Control, indicating that Mn plays an important role in minimizing proton leak dependent respiration. Manganese, through the enzyme Mn Superoxide Dismutase is known to reduce reactive oxygen species in the mitochondrial matrix and reduce proton leak12. Higher State 4 respiration was associated with lower milk and milk protein yield. Since proton leak is an important component of energy efficiency, reducing State 4 respiration through Mn supplementation could improve efficiency.
| Treatments1 | ||||||
| High | Med | Low | LowMn | Control | SEM | |
| Milk, kg | 47.4ab | 50.9a | 46.0ab | 43.6b | 49.7a | 2.9 |
| Milk protein, kg | 1.38ab | 1.44a | 1.40ab | 1.23b | 1.43a | 0.09 |
| State 3 | 75.8 | 64.4 | 78.2 | 73 | 64.1 | 13 |
| State 4 | 26.2ab | 22.6ab | 25.9ab | 27.1a | 22.0b | 3 |
| RCR | 2.89 | 2.76 | 2.98 | 2.65 | 2.83 | 0.27 |
| a b Means within a row not followed by the same superscript letter are significantly different (P < 0.1). | ||||||
| 1 High treatment contains highest levels of Cu, Zn and Mn all well above requirements13, Med treatment contains intermediate levels of Cu, Zn and Mn above requirements, Low treatment contains lower levels of Cu, Zn and Mn but still above requirements, Low Mn treatment contains the lowest levels of Mn (and lower levels of Cu and Zn) but still above requirements and Control treatment contains the lowest levels of Cu and Zn, which are close to requirements. | ||||||
Table 1: Effect of Cu, Mn and Zn supplementation on liver mitochondrial oxygen consumption and milk production from dairy cows at 70 days in milk. This table has been adapted from Acetoze et al. 20177.
Mitochondrial proton leak is a process that dissipates MMP through the movement of protons across the mitochondrial inner membrane without production of ATP14. Proton leak kinetics are assessed by calculating rates of oxygen consumption at a common membrane potential of 165 mV. A lower membrane potential means that protons are 'leaking' across the mitochondrial membrane, which results in lower ATP synthesis (Figure 15). In the Holstein cow study, hepatic proton leak dependent respiration was greatest in LowMn and lowest in Control, which agrees with results in Table 1, that State 4 respiration was greatest in LowMn and lowest in Control.
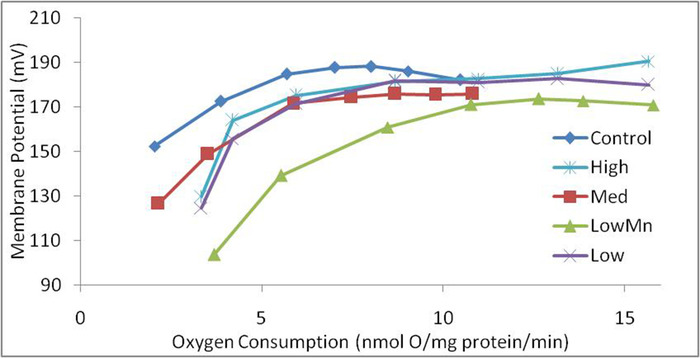
Figure 15. Proton leak kinetics in Holstein cows fed different amounts of Cu, Mn and Zn. This graph is based on data from Acetoze et al. 20177. Please click here to view a larger version of this figure.
Negative results are illustrated in Table 2 and Figure 16. Feed efficiency (RFI) was higher in Angus steers born from low RFI bulls than high RFI bulls, but this was not reflected in mitochondrial RCR (Table 2) or proton leak kinetics (Figure 16). There were no differences in mitochondrial respiration and proton leak kinetics between groups of steers but there was a difference in RFI. There were also no differences (p = 0.88) in hepatic mitochondrial proton leak in high and low RFI steers (Figure 16). There were large standard errors associated with mitochondrial respiration measurements, and proton leak kinetic curves were flat. Liver samples from this study were obtained after steers were slaughtered, a process that delayed liver sample collection and processing by an hour. Variation in mitochondrial respiration measures may reflect mitochondrial respiration degradation due to tissue death. Proton leak kinetic lines were flat because oxygen consumption measurements did not begin until 8 min when plateau had already been reached due to an equipment malfunction.
| Low RFI | High RFI | SEM | P Value | |
| (n=7) | (n=8) | |||
| RFI | -0.58 | -0.01 | 0.1 | 0.05 |
| State 3 | 31.3 | 30.8 | 9.42 | 0.9 |
| State 4 | 9.76 | 10.4 | 3.23 | 0.8 |
| RCR | 3.05 | 3.03 | 0.24 | 0.93 |
Table 2: Performance and mitochondrial respiration of high and low Residual Feed Intake (RFI) Angus bull progeny. This table has been adapted from Acetoze et al. 201511.
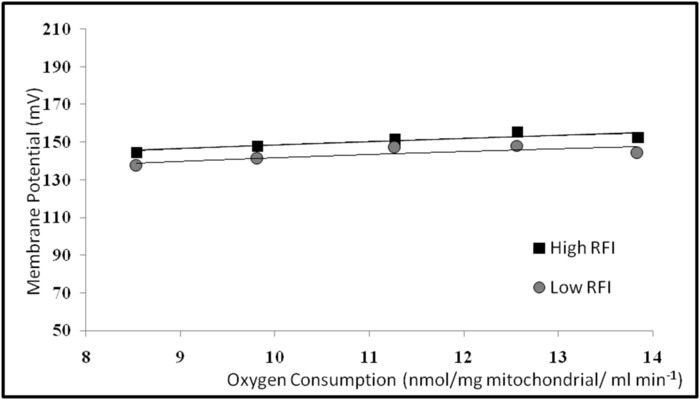
Figure 16. Proton leak kinetics for progeny of high and low RFI Angus bulls. This graph has been adapted from Acetoze et al. 201511. Please click here to view a larger version of this figure.

Figure 1: Clean area for surgical and biopsy materials located in the back of a vehicle and outside of the cow pen. Please click here to view a larger version of this figure.

Figure 2: Restraint of the cow using a halter tied to a cross pole of the head lock. Please click here to view a larger version of this figure.

Figure 3: The area of the cow to clean for the biopsy and location of biopsy at the right 10 – 11th intercostal space found by drawing a straight line from the right tuber coxae to the point of the right shoulder. The biopsy site is where this line intersects with the 10-11th intercostal space. Please click here to view a larger version of this figure.

Figure 4: Shaving a 10 cm area of the cow to prepare to sterilize for the biopsy. Please click here to view a larger version of this figure.

Figure 5: Wash biopsy area of the cow with 10% providone scrub using a circular motion. Please click here to view a larger version of this figure.

Figure 6. Spray biopsy area area with 70% ethanol solution. Please click here to view a larger version of this figure.

Figure 7: Inject 2% lidocaine HCl (10-15 mL) locally to the area to provide anesthesia of the skin. Please click here to view a larger version of this figure.

Figure 8: A 1-2 cm stab-incision through the skin of the 10-11th intercostal space to insert biopsy tool. Please click here to view a larger version of this figure.
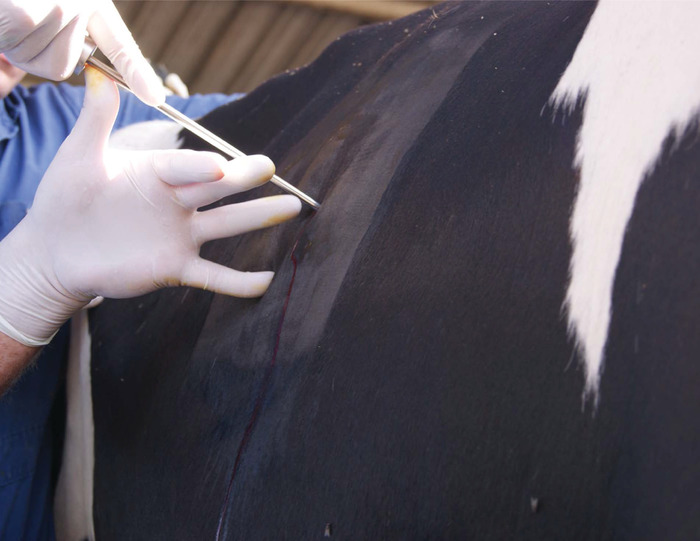
Figure 9: Insertion of bovine liver biopsy instrument through the skin. Please click here to view a larger version of this figure.
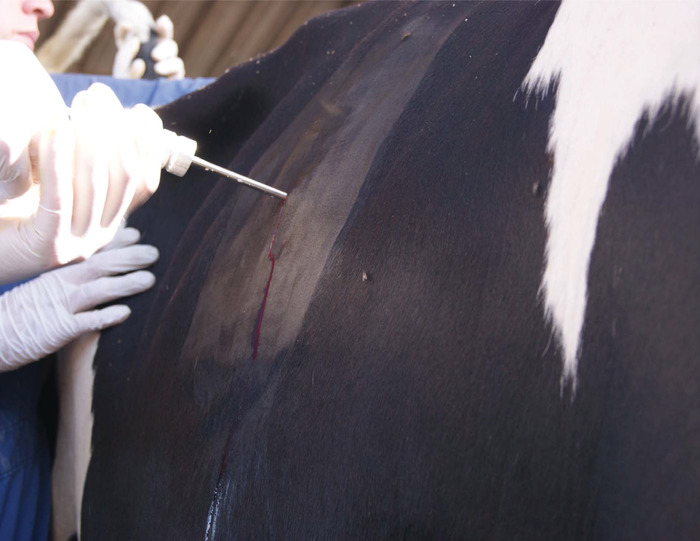
Figure 10: The biopsy instrument should be directed in a slight cranial direction while continuing through the diaphragm and into the liver. Please click here to view a larger version of this figure.

Figure 11: A 1 g sample of the liver being moved from biopsy instrument to Falconer tube for transport to mitochondrial isolation station. Please click here to view a larger version of this figure.

Figure 12: Suturing the skin to close biopsy incision. Please click here to view a larger version of this figure.

Figure 13: Injection of the cow with Ceftiofur Hydrochloride 0.044 mL/kg bodyweight subcutaneously in the neck. Please click here to view a larger version of this figure.
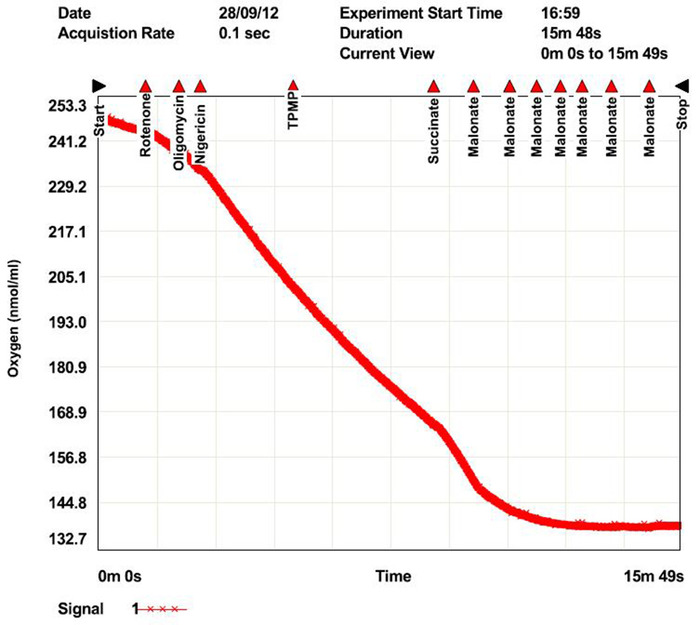
Figure 14: Oxygraph software results showing oxygen consumption responses to addition of each substance to measure mitochondrial membrane potential (MMP) and proton motive force (PMF). Please click here to view a larger version of this figure.
