We designed a whole cell biosensor specific to 2’-FL that can be used in conjunction with the biotechnological production of the oligosaccharide. This relies on the specific enzymatic cleavage of modifying terminal sugars generating lactose and thereby activation of the lac operon, leading to expression of a reporter fluorescent protein under a lactose inducible promoter, in proportion to the quantity of 2’-FL. To demonstrate its linkage specificity, 3-fucosyllactose (3-FL), an isomer, was also tested. The genetic circuit contains two parts: a fucosidase gene, afcA or afcB, driven by a constitutive promoter resulting in constitutive expression cloned on to a vector with a signal sequence of pelB encoded upstream of the fucosidase gene to facilitate periplasmic export, as well as a fluorescent reporter system containing the T7 promoter driving expression of GFP. The first part is expressed on plasmid pAfcA and the second reporter system on plasmid pET28:GFP. The final construct was transformed into E. coli BL21 (DE3), a widely available strain that has the T7 RNA polymerase integrated into the genome under control of the lactose-responsive pLacUV5 promoter. Figure 1A shows the schematic design so that the reader can follow the working principle of the biosensor. Figure 1B illustrates the fluorescent signal under different conditions of induction as analyzed on a flow cytometer. Consistent with our hypothesis, over a 100-fold increase in fluorescence was observed with the 2’-FL biosensor compared to the vector-only control or with the biosensor for 3-FL. This shows the high linkage-specificity of the biosensor. To ensure that the signal is indeed due to defucosylation, we ran a control by adding commercial α-1,2-fucosidase to the culture during induction, which is confirmed as shown by the bars on the right. The sensitivity of the assay can be determined by generating a dose-response curve with varying carbohydrate levels (Figure 2A). From the plot, the dynamic linear range of 2’-FL detection is between 40 and 400 mg/L and the limit of detection is 4 mg/L. This assay can be carried out over the course of a working day, as shown by the snapshot measurements of the dynamics of reporter expression (Figure 2B).
Finally, we present how the biosensing cells can be used to detect and quantify 2’-FL from an engineered 2’-FL producer strain. Since this strain uses lactose as a substrate, we developed strategies to reduce signal from exogenous lactose so that the signal readout would directly correspond to the 2’-FL concentration (Figure 3A). One strategy is to enzymatically degrade the lactose using a β-galactosidase that does not act on modified lactose. This can be achieved using commercial β-galactosidase that preferentially acts on lactose, reducing the lactose signal to 20% of the original (Figure 3B) or via incubation with a LacZ+ strain, wild-type BL21 (DE3), which in three hours drops the lactose signal to background levels (Figure 3C). The last strategy utilizes ethanol, which not only selectively precipitates lactose but also lyses the producer cells, cell lysis being a necessary step downstream (Figure 3D). This is a simple extraction process, but care should be taken due to lower recovery of 2’-FL compared with other methods, possibly from losses due to some crystallization of 2’-FL. In spite of the addition of ethanol, we did not observe significant inhibition of growth of the biosensing cells, likely because the final ethanol concentrations are low (<2%, v/v) in the cultures. Finally, the most effective but time-consuming method we demonstrate is to pellet and wash the cells prior to cell lysis to release the intracellular 2’-FL (Figure 3E). We advise the reader to use discretion for selecting the appropriate method for lactose removal based on efficiency and availability of time and reagents.
Figure 3E also demonstrates our ability to reliably quantify 2’-FL in biotechnological context. We cultured either an engineered 2’-FL-producing strain11 or a fucosyltransferase negative mutant (negative control) with lactose to monitor production of 2’-FL in cell lysate. After culturing the cells, we measured the concentration using both the biosensor and high-performance liquid chromatography (HPLC) for validation (Supplementary Figure 1). The dose response curves for producer lysate and control lysate spiked with 2’-FL are very similar, which demonstrates quantitative measurement of 2’-FL in the context of cell lysate.
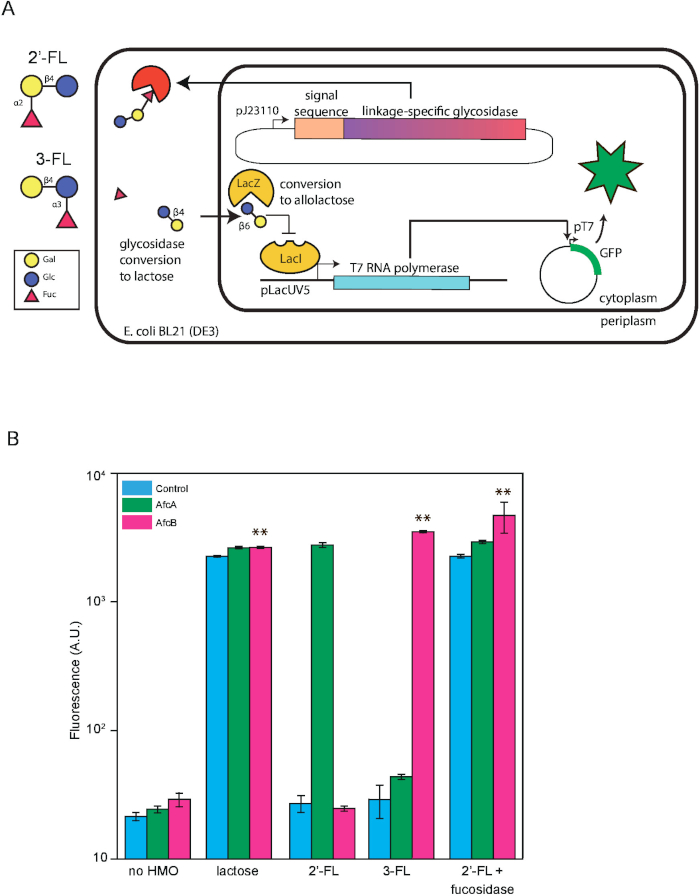
Figure 1: Schematic of 2’-FL biosensor design and representative data. (A) 2’-Fucosyllactose (2’-FL) enters the bacterial periplasm and is converted to lactose by heterologously expressed fucosidase. Lactose is transported to the cytoplasm and converted to allolactose. This activates the native LacUV5 promoter in E. coli BL21 (DE3), producing T7 RNAP, which transcribes GFP under the T7 promoter, leading to fluorescent protein expression. (B) Detection of 2’-FL and 3-FL. Cells containing pET28:GFP and either pAfcA (green), pAfcB (magenta), or an empty vector control (blue) were induced overnight with no sugar, 2’-FL, 2’-FL with exogenously added α-1,2-fucosidase, 3-FL, or lactose. All data are an average of three biological replicates each analyzed in triplicate, where error bars represent standard deviation from the mean. Statistical significance was determined by Tukey’s multiple comparison test and ** is indicative of p < 0.01. This figure first appeared in Enam and Mansell14 and is reused with permission. Please click here to view a larger version of this figure.
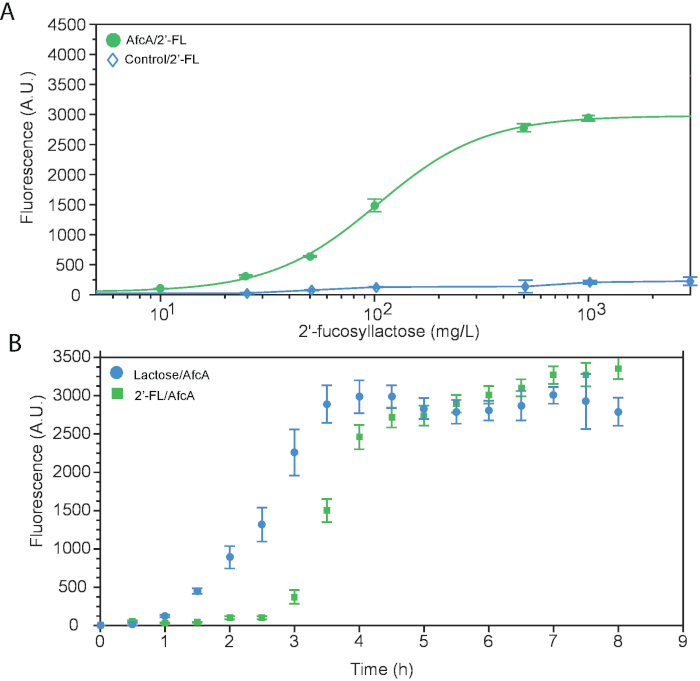
Figure 2: Characterization of biosensor for 2’-FL detection. (A) Limits of detection and transfer functions of biosensors. Response curves depicting mean fluorescence for pAfcA/2’FL (green circle) or empty control vector (blue diamond) at varying amounts of 2’-FL cultured. (B) Time course of reporter expression. Strain pAfcA was incubated with 2’-FL (green square) and fluorescence was monitored over 8 h of induction. Strain pAfcA incubated with lactose is included as a control. All data are an average of three biological replicates each analyzed in triplicate, where error bars represent standard deviation from the mean. This figure first appeared in Enam and Mansell14 and is reused with permission. Please click here to view a larger version of this figure.
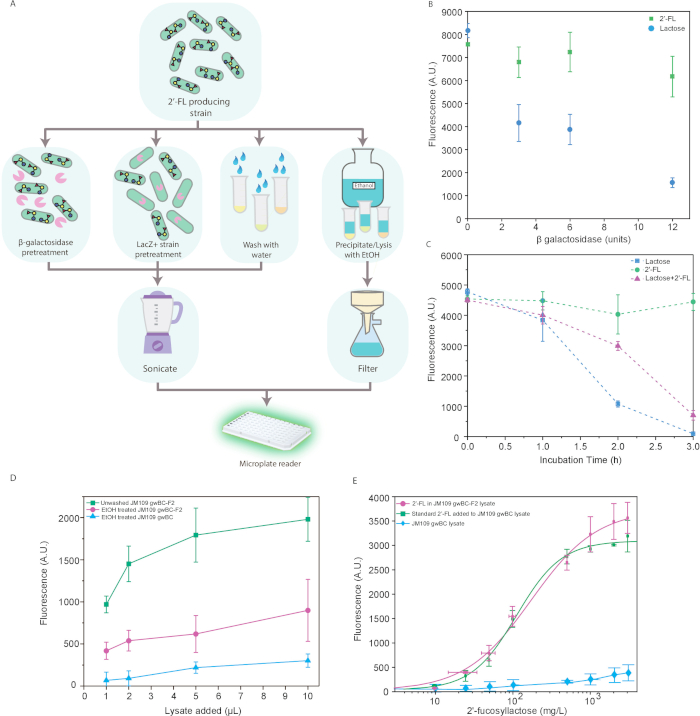
Figure 3: Detection and quantification of 2’-FL from a producer strain. (A) Workflow demonstrating the use of the biosensor in a simple protocol with key focus on removing noise from residual lactose. (B) Removal of lactose by β-galactosidase enzymatic digestion. Cultures containing 0.3% 2’-FL (green square) or 0.2 % lactose (blue circle) were incubated with biosensor cultures containing varying amounts of exogenous β-galactosidase at induction time were assayed for fluorescence after incubation overnight. (C) Removal of lactose by incubation with wild-type LacZ+ strain. LB containing 0.3% 2’-FL (green circle), 0.2% lactose (blue square), or a 1:1 mixture (0.2% lactose equivalent, i.e., 0.1% lactose + 0.15% 2'-FL, magenta triangle) was incubated for various times with BL21 (DE3) cells, and then added to biosensor cultures for fluorescence measurement after incubation overnight. (D) Precipitation of lactose and cell lysis by ethanol pretreatment. Fermentations of JM109 gwBC-F2 (magenta circle) or JM109 gwBC (blue triangle) cells were incubated with 2 volumes of ethanol and filtered before incubation with fluorescent biosensor cells and compared with fluorescence from untreated JM109 gwBC-F2 culture (green square). (E) Quantification of 2’-FL in cell lysate. The biosensor was evaluated for its ability to detect 2’-FL from a metabolically engineered producer strain. Fluorescence measurements generated by addition of either crude lysate of JM109 gwBC-F2 (magenta circle, 2’-FL as quantified by LC-MS), 2’-FL in defined amounts to nonproducer strain JM109 gwBC cell lysate (green square), or crude lysate of JM109 gwBC strain (blue diamond). Results were validated by HPLC quantification of 2’-FL in producer strain. All data are an average of three biological replicates each analyzed in triplicate, where vertical error bars represent standard deviation from the mean fluorescence and horizontal error bars represent the standard deviation in 2’-FL measured by HPLC in the triplicates. This figure first appeared in Enam and Mansell14 and is reused with permission. Please click here to view a larger version of this figure.
Supplemental Figure 1: HPLC-MS analysis of 2’-FL. (A) Extracted ion chromatograms of 2’-FL (m/z 511). Chromatogram of a commercial standard is shown on the top. Chromatogram of 2’-FL from the JM109gwBC-F2 strain (10-fold dilution) is shown at the bottom. The peak at 511 m/z is the sodium adduct. (B) An enlargement of mass spectrum m/z from 300−800, where the most abundant signals were concentrated is shown for the commercial standard (top) and the 2’-FL from the producer strain (bottom). (C) A standard curve was created by LC-MS analysis with varying levels of commercial 2’-FL. Error bars represent standard deviations from triplicate measurements. Please click here to download this file.
